De appelvorm versus de peervorm
De eerste aanwijzing dat je misschien insuline resistent bent, is de verdeling van het vet op je lichaam, zoals gedemonstreerd in de “appel” vs “peer”-vormige beelden van het lichaam.
Insulineresistentie wordt gekenmerkt door abdominale vet, in de “appel” vorm aan de linkerkant. De rollen van vet rond de buik, de gezwollen buik – dat zijn de klassieke tekenen. De ophoping van vet rond en boven de taille is een gevaar teken.
In de “peer” vorm, het gewicht is meestal lager dan de taille, op de heupen en dijen, en de buik is duidelijk vlakker. Als je peervormig, bent u waarschijnlijk tot insulineresistentie te hebben.
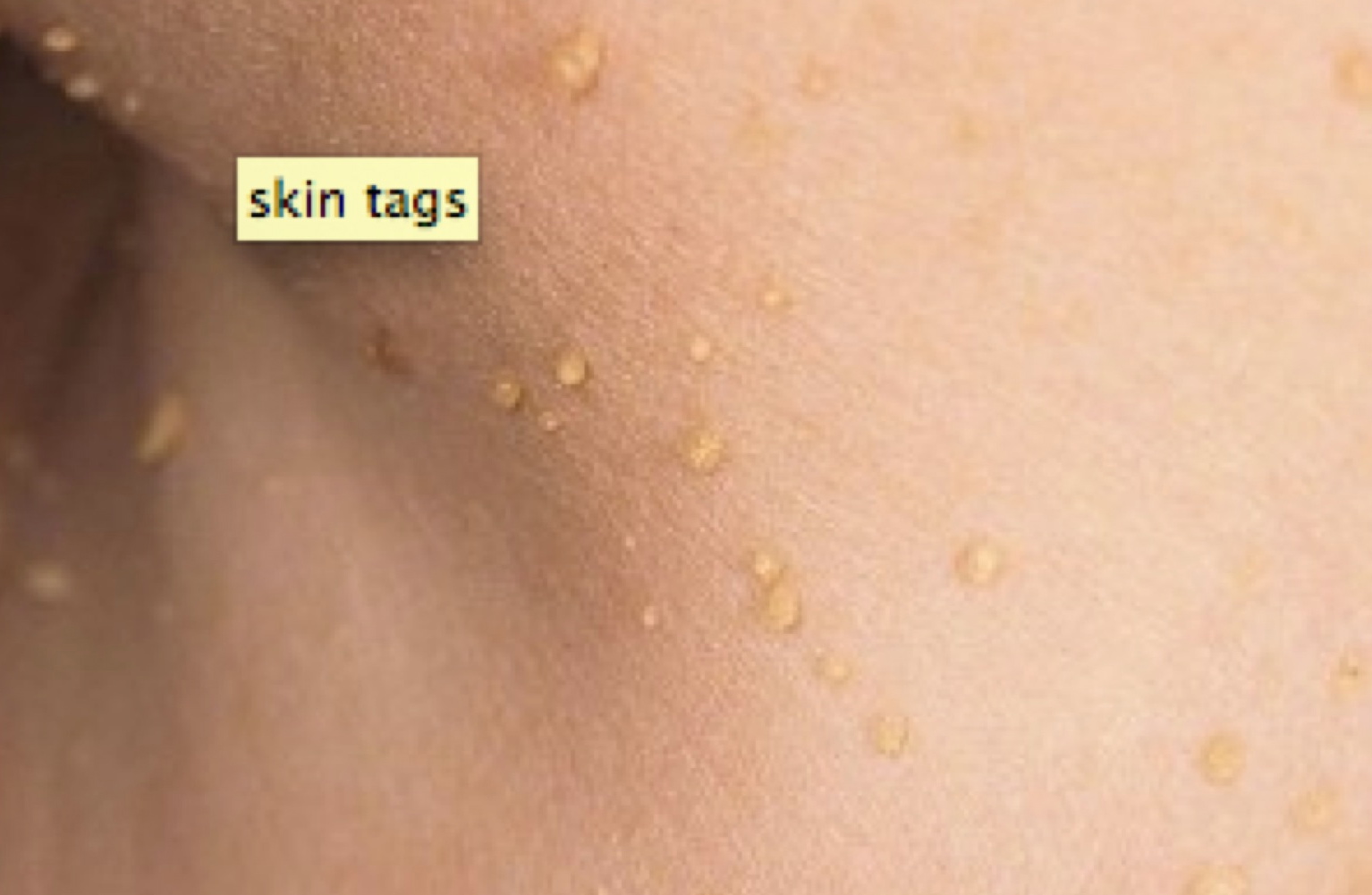
Skin tags
Een ander symptoom dat is bijna altijd een teken van insulineresistentie zijn de huid tags. Skin tags zijn kleine bultjes of knobbeltjes op de huid, die aanwezig kunnen zijn op de hals, gezicht, in de oksels of op andere plaatsen waar de huid is gekreukt.
Let op: De aanwezigheid van de huid tags is een zeer sterke indicator dat u insuline resistentie hebben.
Acanthosis nigricans
Acanthosis nigricans is de technische term voor een donkere vlek van de huid. Het wordt vaak gezien rond de hals en onder de armen, op ellebogen, knieën en knokkels. Nogmaals, het is niet gevaarlijk, maar als je dit symptoom hebt, dan bent u zeer waarschijnlijk insuline resistent.
Niet helemaal in goed Nederland’s vertaald, maar evengoed heeft AW de moeite voor u gedaan om het enigszins begrijpelijk te vertalen.
Meer dan enkele honderden miljoenen mensen zullen als uitings vorm diabetes hebben en obesitas zijn in de komende decennia, terwijl de werkelijke therapeutische benaderingen gericht op het behandelen van de gevolgen in plaats van de oorzaken te achterhalen door de stofwisseling te verminderen. Deze strategie is niet efficiënt en nieuwe paradigma’s moeten worden gevonden. De brede analyse van het genoom kan niet voorspellen of verklaren 10-20% van de ziektemeer dan , terwijl veranderingen in de voeding en sociaal gedrag hebben zeker een grote impact. Echter, de moleculaire mechanismen die omgevingsfactoren en genetische gevoeligheid waren tot nu toe niet voor ogen tot aan de recente ontdekking van een verborgen bron van genomische diversiteit, dat wil zeggen, de metagenoom. Meer dan 3 miljoen genen van enkele honderden soorten vormen onze intestinale microbioom. Eerste toets experimenten hebben aangetoond dat dit biome uit zichzelf stofwisselingsziekte brengen. De mechanismen zijn onbekend maar kunnen worden betrokken in de modulatie van energie vangstcapaciteit door de gastheer en de laagwaardige ontsteking en de overeenkomstige immuunrespons op vetweefsel plasticiteit, hepatische insulineresistentie en zelfs de secundaire cardiovasculaire gebeurtenissen. Uitgescheiden bacteriële factoren bij de circulerende bloed, en zelfs volledige bacteriën uit darmflora kan bereiken weefsels waar ontstekingen wordt geactiveerd. De laatste 5 jaar hebben aangetoond dat darmflora, op zijn moleculair niveau een oorzakelijke factor in de vroege ontwikkeling van de ziekten. Toch veel meer behoefte worden blootgelegd om eerste, nieuwe voorspellende biomarkers te identificeren, zodat preventieve strategieën op basis van pre-en probiotica, en ten tweede, nieuwe therapeutische strategieën tegen de oorzaak in plaats van het gevolg van hyperglycemie en gewichtstoename.
introductie
Metabole ziekten zoals diabetes en obesitas worden een sociaal probleem van het grootste belang voor alle landen. Hun impact op ontwikkelingslanden zoals Zuid-Azië is nog dramatischer omdat, naast wordt beïnvloed door de hoogste groeiende percentage, het sociale systeem kan zeker niet veroorloven de bijbehorende kosten. Daarom wordt de ziekte slecht behandeld en pathologische complicaties staan in bloei. Inderdaad een belangrijk resultaat gekoppeld aan het voorkomen van metabole ziekten is de snelle toename van cardiovasculaire gebeurtenissen die leiden tot de dood [1]. In de afgelopen tien jaar is diabetes de oorzaak geweest van dodelijke cardiovasculaire gebeurtenissen die zijn gevorderd de meeste, aangezien een stijging met 62% is gekwantificeerd [2, 3]. De progressie is veel hoger dan het risico toegewezen aan cholesterol of hypertensie. Derhalve in westerse landen, waar stofwisselingsziekten worden geïnstalleerd, en in Oosten, waar diabetes en obesitas sterk in opkomst is er een dringende behoefte om eerst risicofactoren van diabetes en obesitas en tweede nieuwe therapeutische targets te identificeren. Bijgevolg preventieve strategieën, waarschijnlijk op basis van een verandering in eetgewoonten of het gebruik van voedsel vormt een aanvulling op prebiotische en probiotische, massaal zou kunnen worden gelanceerd. Ten tweede is therapeutische behandelingen die gericht zijn op behandeling van de oorzaak niet het gevolg van een verhoogde lichaamsgewicht of glycemie nodig. Om dit doel is een nieuw paradigma van metabole problemen nu in opkomst, wat resulteert in de herziening van de onderliggende oorzakelijke factoren.
Diabetes en obesitas het gevolg zijn van genetische en omgevingsfactoren. Geschat wordt dat puntmutaties voor minder dan 10% van de totale fenotype. De lage impact van genetica op metabole ziekten wordt nog versterkt door de toenemende incidentie van diabetes en obesitas in de afgelopen decennia. De incidentie van type 2 diabetes bereikt 4-5% in Europa, 8-10% in de VS en meer in Zuid-Azië [4]. Deze aantallen zijn meer dan verdubbeld in de afgelopen 20 jaar. Derhalve kan suggereren dat zelfs als genetische analyses de basis voor deze epidemie veranderingen in ons genoom niet alleen verantwoordelijk [5]. Talrijke andere hypothesen zijn voorgesteld. Ten eerste, epigenetische niet-gecodeerde factoren genereren van een nieuwe veelbelovende tijdperk van hypothesen die zou moeten worden bestudeerd en dat niet afhankelijk zou zijn van genomische spelers. Ten tweede, realistischer is de impact van een verandering in de eetgewoonten en het sociale gedrag dat zijn zeker belangrijke oorzaken van de toenemende incidentie van metabole ziekten. Echter, beide factoren immers niet uit algemene epidemie. Gekoppeld aan de milieu-hypothese, hebben recente bewijzen bracht de metagenoom hypothese. De laatste wordt gedefinieerd als de totale bacteriële genoom, terwijl de expressie van het overeenkomstige gen vertegenwoordigt de metatranscriptome. Deze prokaryote genoom, naast onze eukaryotische een, ontstaan door recente ontwikkelingen van high throughput sequencing technologieën voor miljoenen verkrijgen leest gedurende een korte tijd en vermeden dat laboratorium kweek van darmbacteriën [6], zelfs als een recent studie lijkt dit concept achterwaarts [7]. De recente ontwikkelingen in de DNA-sequencing technologie hebben toegelaten dat de verzameling van hoog-dimensionale data van menselijk-geassocieerde microbiële gemeenschappen op een ongekende schaal [8]. Daarnaast zijn enorme inspanningen geleverd in bioinformatische analyse waardoor het coderen en het decoderen van alle sequenties. Mensen gastheer verschillende metagenomes van meerdere locaties zoals huid, longen, vagina, mond, zelfs als de gastheer de darm [9, 10]. In feite is de menselijke darm 100000000000000 gastheer micro-organismen omvat tot duizenden soorten in een gemiddelde concentratie van 1014 per ml en een gewicht gemiddeld 1,5 kg [11]. Het belang van deze metagenoom ligt in haar genen repertoire, 100 keer beter dan onze eukaryote nucleaire genoom [12], die dus een grote genetische diversiteit gevoelig voor een enorme hoeveelheid functies over te brengen.
Een belangrijk concept is dat alle zoogdieren zijn steriel geboren, zonder enige flora. Na de eerste uren, dagen en weken van de moeder en de milieu-flora gekoloniseerd het gehele lichaam van de nieuwe geboren in een bepaalde volgorde [13]. De initiële kind darmflora is een eenvoudige structuur gewoonlijk beheerst door Bifidobacteria, en door een reeks opeenvolgingen en vervangingen, verschuift een complexer patroon volwassene [14-16]. De microbiota ondergaat ook inhoudelijke veranderingen in de uitersten van het leven, bij zuigelingen en oudere mensen, de gevolgen van die nog steeds worden onderzocht [17]. Echter, de volwassen intestinale microbiota is aangetoond dat relatief stabiel zijn over de tijd [18] en voldoende gelijkenis tussen individuen van de identificatie van een kern Microbiome bestaande uit 66 dominante operationele taxonomische eenheden (Otus) die overeenkomt met 38% van de volgorde leest van 17 personen [19]. Uiteindelijk, tot de kern microbiota veranderingen worden onderscheiden bij oudere patiënten van die waargenomen voor jongere volwassenen, met een groter deel van Bacteroides spp. en onderscheiden overvloed patronen van Clostridium groepen. Ecologische regels gelden voor de vorm van de microbiële diversiteit gedurende het hele leven. Dit suggereert dat elk lid kan deelnemen aan een perfect mutualistische symbiose met elkaar en definieert een constante microbiota [9, 10, 20]. Echter, voordat een ideale microbiële ecologie de microben interactie met de host. De interactie van epitheelcellen met bacteriën en componenten vrijgegeven door microben, met inbegrip van de metabolieten, is een belangrijke mediator van de overspraak tussen epitheel en andere celtypen [21]. De uitwisseling tussen bacteriën en epithelium kunnen verschillen in de dunne en dikke darm door anatomische verschillen en de mate waarin de uitgescheiden slijm bedekt de epitheel. Een belangrijk kenmerk is dat de mucosa is vrij van bacteriën, echter dit niet uit dat bacteriële deeltjes kan diffunderen in de mucosa aan receptoren binden aan het oppervlak van epitheliale darmcellen [22]. Deze bacterie om interactie gastheer helpt rijp worden de intestinale epitheliale laag, de mucosale aangeboren immuunsysteem, het enterale zenuwstelsel, evenals de darm vasculaire systeem [21]. Vandaar darmflora heeft een sterke invloed op de beheersing van de talrijke belangrijke fysiologische functies. Of de eerste jaren van het leven van invloed kan zijn volwassen fysiologische functies wordt vermoed. Een interessante kanttekening is dat de moderne normen van hygiëne [23] en / of de overstap van borstvoeding naar zuigfles kan transmissiemechanismen [13, 24] hebben veranderd. Vergelijkbare hypothesen zou kunnen worden opgetrokken ten aanzien van premature neonaten [25]. Al met al is het denkbaar dat de vroege intestinaal kolonisatie bij de geboorte invloed kan het optreden van metabole ziekten [26]. Recente cijfers dat de Bifido bacteriële telling in fecale monsters tijdens de kinderschoenen staat als, beoordeeld door FISH met flowcytometrie, was hoger bij kinderen nog een normaal gewicht over een 7-jaar follow-up studie [26]. Omgekeerd werd de microbiota dwaling bij zuigelingen in kinderen aan overgewicht geassocieerd met een groter aantal Staphylococcus aureus.
Eenmaal volwassen microbiota gevestigd, kon actoren zoals antibiotica, prebiotica en probiotica moduleren zijn ecologische architectuur. Echter, hoewel sommige plasticiteit bestaat, deze effecten zijn altijd omkeerbaar, wat suggereert dat een strakke host-naar-microbiota relatie is vastgesteld tijdens de neonatale leven waar de gastheer geeft vorm aan de microbiota en vice versa [27]. De overeenkomstige mechanismen kan worden gekoppeld aan de rijping van het immuunsysteem [28] waarbij sommige bacteriële species zoals gesegmenteerde filamenteuze bacteriën grotendeels kunnen de gecoördineerde rijping van T-cellen, waarvan de reacties worden geïnduceerd door de gehele microbiota induceren. Het aangeboren immuunsysteem, als een eerste lijn van verdediging, houdt steriel de intestinale mucosale laag door de fagocytose van binnendringende ziekteverwekkende bacteriën. Secundair, zal B-lymfocyten scheiden IgA in de mucosa die specifiek zullen worden gericht tegen invasieve pathogenen [29 tot 31]. Uiteindelijk zal de synthese van darmepitheelcellen van lectine voorzien strooisel voor commensale bacteriën mucosale en de secretie van defensine regelen. De wijze waarop deze twee, natuurlijke antibiotica uitgescheiden door de Paneth cellen, selecteert sommige bacteriële geslachten en soorten die zullen blijven dicht bij het darmslijmvlies.
Rekening houdend met alle bovenstaande opties, blijft het bepalen van de belangrijkste factoren die verantwoordelijk zijn voor de epidemie van metabole ziekten die snel ontgroeien. Onlangs heeft darmflora ecologie aangetoond metabole ziekten zoals obesitas zelf [32] en diabetes vorm [33]. High throughput sequencing analyses toonden een verandering in enkele grote phyla die in deze beoordeling worden beschreven volgens het fenotype. Bij mensen, een grote vraag blijft of darmflora het gevolg of de oorzaak van de waargenomen fenotype. Hoewel studies met kiemvrije muizen aangetoond dat de causale rol van intestinale microbiota bij het ontstaan van metabole stoornissen, blijft het nog steeds aan te tonen of de genetische achtergrond kan de ontwikkeling van een specifieke microbiota beïnvloeden.
Dat het dieet kan een belangrijke regulator factor in het vormgeven van darmflora en daarmee de relatie met gastheer stofwisseling, logisch kan worden aangenomen, gezien de belangrijke rol van de darmflora in de spijsvertering [34]. In feite is een vergelijkende studie van verschillende voedingsgewoonten, zoals herbivoor, omnivoor en carnivoor aangetoond dat het verwerven van een nieuw dieet is voldoende om radicaal te wijzigen darmflora, als een evolutionaire oorzaak van nieuwe soorten [35]. In feite, zowel gastheer dieet en fylogenie vorm bacteriële variëteit, die toeneemt van vlees naar herbivoor naar omnivoor. Bovendien een moderne levensstijl verschuift darmflora van mensen dichter bij die van omnivore primaten [35]. Daarom is het vanzelfsprekend dat onze darmflora voedt zich met het soort voedsel dat wordt geabsorbeerd door het veranderen van de ecologische structuur. Elke individuele bacterie leeft in mutualistische ecologie met de anderen. Daarom kan een overmaat of een gebrek aan nutriënten veranderen de metabolische activiteit van een bepaalde bacterie, die niet meer of uiterst producten zoals een metaboliet essentieel of schadelijk voor de naburige bacteriën. De belangrijkste voorbeeld is zuurstof die wordt gebruikt door de aeroben van het bovenste maag-darmkanaal het induceren van een diepe anaërobe toestand in de distale darm waardoor dus strikt anaëroben om te overleven. Het gebruik van prebiotica zoals niet-verteerbare polysacchariden dieet als substraten voor genera zoals Bifidobacterium zullen bevorderen de groei en het anti-inflammatoire functie [36]. Ook een vet-verrijkt dieet, die op grote schaal wordt gebruikt om metabole ziekten veroorzaken, van grote invloed op de ontwikkeling van diabetes en obesitas.
De moleculaire mechanismen waarmee een bepaalde darmflora induceert metabole ziekten zal worden besproken in deze review. Kort ze zijn gekoppeld aan een verhoogde energie oogsten en het ontstaan van de low-grade inflammatoire status karakteriseren insulineresistentie en obesitas [37, 38]. De bacteriële moleculen die verantwoordelijk zijn voor de activering van stellingen fysiologische functies nu ontdekt. Dit zal zeker leiden tot de identificatie van nieuwe therapeutische strategieën voor de behandeling, maar ook voor het voorkomen van metabole ziekten. Een tweede uitkomst zal de identificatie van biomarkers in staat om de ontwikkeling van diabetes en overgewicht in de afwezigheid van een risicofactor te voorspellen zijn. Dit wordt voorzien aangezien darmflora causaal is voor de ontwikkeling van metabole ziekten.
Al met al, de nieuwe “microbiota aan paradigma host” voor de controle van metabole ziekten is veelbelovend. Het gaat om transversale gebied van onderzoek, zoals microbiologie, immunologie, metabolisme, en bio-informatica. Deelnemen aan de verschillende stukjes van de puzzel moet leiden tot innovatieve preventieve en therapeutische strategieën die farmacologische, prebiotische en probiotische ontwikkelingen en synbiotische aanpak [39] te betrekken.
Veranderingen in microbiota tijdens metabole ziekten
In de afgelopen ~ 160 miljoen jaar, hebben zoogdieren samen geëvolueerd met een grote en diverse microbiële gemeenschap die onze huid en mucosale oppervlakken koloniseert. De meeste van deze organismen bevinden in ons maagdarmkanaal en aanleiding geeft wordt bepaald door gastheer fylogenie en dieet [40, 41]. Aangezien 0,3% van eukaryotische genoom is gemodificeerd 1 miljoen jaar kan worden berekend dat 50-55% van ons genoom geëvolueerd in een strakke relatie met de microbiële gemeenschap die sterke genetische afhankelijkheid. Naast deze zeer lange termijn genetische evolutie, de afgelopen tien jaar, is een belangrijke constatering is dat metabole ziekten gaan gepaard met een verandering in de darmflora samenstelling bij dieren en mensen [32, 42, 43]. De ontwikkeling van high-throughput sequencing technologieën, zoals pyrosequencing, heeft het mogelijk gemaakt de analyse van de uitwerpselen van patiënten met obesitas tijdens gewichtsverlies. Kortom, deze technologie sequenties miljoenen 16S rRNA genfragmenten per run. Het 16S rRNA uit 9 hyper-variabele gebieden (V1-V9) [44, 45] is de doelstelling voor de amplificatie gebaseerde sequencing vertegenwoordigen. De mate van diversiteit of homologie is evenredig met de genetische afstand wordt uitgezet op een fylogenetische boom. Vandaar dat deze kenmerken leiden tot de identificatie van bacteriële Phyla, klassen, orden, families, geslachten en soorten in vergelijking met een naïeve data basis [46]. Hoe langer de sequentie des te nauwkeuriger wordt de analyse. Het gebruik van merktekens verschillende monsters te identificeren kan de analyse van meerdere monsters tegelijkertijd, waardoor dus de invloed van sequentie verschillen in effectiviteit. Echter, deze strategie vermindert ook de diepte van de analyse door het aantal leesbewerkingen per monster. De volgorde leest zijn toegewezen aan de NCBI niet-overtollige, Clusters van orthologe groepen (COG) [47] of Kyoto Encyclopedia of genen en genomen (KEGG) [48] databases. Totaal in het algemeen wordt, zijn de gemiddelde resultaten van alle datasets gerapporteerd als volgt: 94% van de labels die aan de niet-redundante databank werden bacteriële, eukaryote waren 3,6% (0,29% Mus musculus, 0,36% schimmel) 1,5% waren Archaea (1,4% Euryarcheota; 0,07% Crenarcheota), en 0,61% waren virale (0,57% dubbel-strengs DNA-virussen) [49]. Eerste analyses met deze techniek zijn uitgevoerd op obese patiënten tot gedurende 1 jaar beperking calorieën diëten gevolgd om hun lichaamsgewicht te verminderen. Gordon groep aanvankelijk aangetoond dat de obese patiënten gekenmerkt worden door veranderingen in de relatieve hoeveelheden van de twee dominante bacteriële divisies de Bacteroidetes en firmicutes [32, 50]. Zij toonden een verminderde Bacteriodetes / firmicutes verhouding bij obese patiënten die in de richting van die van magere patiënten geëvolueerd tijdens gewichtsverlies. Aldus wordt geassocieerd met obesitas phylum niveau veranderingen in de microbiota en verminderde bacteriële diversiteit [32, 43, 51]. Daarom werd een verandering in darmflora gekoppeld aan de obese fenotype. Twee hypothesen kan worden verhoogd op basis van deze resultaten. Het obees fenotype zou zijn ondergeschikt aan de microbiota of het tegenovergestelde. Om deze vraag te beantwoorden, hebben de fecale microbiële gemeenschappen van volwassen vrouwelijke eeneiige en twee-eiige tweelingen concordant voor magerheid of obesitas, en hun moeders onderzocht [51]. De resultaten tonen aan dat de menselijke darm microbioom wordt gedeeld door familieleden, maar dat elke persoon darm microbiële gemeenschap varieert in de specifieke bacteriële geslachten aanwezig zijn, met een vergelijkbare mate van co-variatie tussen volwassen eeneiige en twee-eiige tweelingen [51]. Deze resultaten demonstreren dat een diversiteit van organismale assemblages kan echter een kern microbiome op genniveau en afwijkingen van deze kern zijn geassocieerd met verschillende fysiologische toestanden, bijvoorbeeld obese versus lean geven.
Opmerkelijk is dat deze technologie snel ontwikkelt en dat nieuwe generaties sequencing met verschillende bioinformatica analyses waardoor een nog snellere analyse van de microbiome. Deze technieken gebruikt Illumina GAIIx platform sequentie een uiteenlopende reeks van monsters op een diepte van enkele miljoenen middeling leest per sample die continu toeneemt [52]. De gegevens tonen een uitstekende consistentie in taxonomische diversiteit heroveren herstel en patronen die voorheen werden gerapporteerd op basis van meta-analyse van vele studies uit de literatuur. Het gebruik van deze techniek heeft het bestaan van de hierboven gerapporteerde kern microbioom [51].
Recente gegevens gekenmerkt intestinale microbiota in type 2 diabetes patiënten. De auteurs beschreven dat het aandeel phylum firmicutes en klasse Clostridia aanzienlijk werden verlaagd bij diabetici in vergelijking met de controlegroep [33]. Bovendien de verhoudingen van Bacteroidetes te firmicutes en de verhoudingen van Bacteroides-Prevotella groep C. coccoides-E. rectale groep positief en significant gecorreleerd met plasma glucose concentratie, maar niet met een BMI. Daarom kan bacteriële sequenties, specifiek voor type 2 diabetes dan obesitas, geacht handtekeningen van hyperglycemische syndroom.
Oudere gerapporteerde gegevens veranderingen in de darmflora in verband met verschillende metabole fenotypes. Dertig jaar geleden werd waargenomen tijdens de maag-bypass operatie, een chirurgische methode nu op grote schaal uitgevoerd om diabetes en morbide obesitas te behandelen [53], een verandering in de darmflora, zoals waargenomen door de cultuur-gebaseerde methoden [54]. Bovendien in diermodellen van obesitas veroorzaakt door hersenletsel van de ventromediale hypothalamus, een verandering van darmflora werd ook waargenomen suggereert een sterke invloed van de hersenen op de bestrijding van darmflora [55]. Op dat moment, de teelt-gebaseerde technologieën toonde veranderingen in enterocci en lactobacillen.
Het gebruik van metagenomic sequenties moeten nu helpen om nauwkeurig de rol van de hersenen betreffende de controle op microbiota. Volgens dezelfde observatie van de genetische ablatie van de leptine receptor gen in muizen herbergt een microbiota dat een aanzienlijk hoger percentage firmicutes bezit, en een dienovereenkomstig lager percentage Bacteroidetes dan hun wild-type nestgenoten [49]. Deze dataset toont aan dat leptine zeker darmflora regelt. De moleculaire relais koppelen leptine aan de microbioom zijn onbekend, maar de gegevens wijzen erop dat haar optreden zou zijn, zowel via de centrale ingang of via de inductie van obesitas. Bovendien is recent aangetoond een relatie tussen darmflora en hersenontwikkeling sinds kiemvrije muizen tonen verhoogde motorische activiteit en verminderde angst, in vergelijking met specifieke pathogeenvrije (SPF) muizen met een normale darmflora [56]. Met name de auteurs toonden aan dat de microbiële kolonisatie signaleringsmechanismen staat om neuronale circuits betrokken bij motorische controle en angst beïnvloeden control initieert.
Een andere hypothese wordt gekoppeld aan de rol van hormonen op het immuunsysteem dat uit zichzelf kan darmflora vorm zoals voor type 1 diabetes model, waarbij het immuunsysteem is aangetast en in andere gevallen en [57-59]. Inderdaad, een mogelijke uitkomst van de adaptieve co-evolutie van de mens en bacteriën is de ontwikkeling van commensale relatie [60]. Een belangrijk mechanisme omvat de secretie van antimicrobiële moleculen die behoren tot de familie of defensine leden van de familie, zoals RNAse angiogenin [58, 59]. Deze moleculen door intestinale Paneth cellen worden uitgescheiden in de darmlumen en hebben bactericide activiteit tegen intestinale microben. De expressie wordt geïnduceerd door Bacteroides thetaiotaomicron een overheersende lid van de darm microflora. Muizen verwijderd uit de overeenkomstige genen hebben verschillende microbiota, het onthullen van een mechanisme waarbij intestinale commensale bacteriën beïnvloeden darm microbiële ecologie en vorm aangeboren immuniteit [58]. Het immuunsysteem kan dan regelen de secretie van defensine en vormen de microbiële gemeenschap [61]. Daarom zou men kunnen suggereren dat in stofwisselingsziekten, de belangrijke rol van ontsteking [37, 38, 62], vooral als gevolg van een verzwakt immuunsysteem, anders zou darmflora vorm te geven.
Voeding en voedingstoestand behoren tot de belangrijkste beïnvloedbare factoren voor de menselijke gezondheid. Bovendien een verandering van eetgewoonten is waarschijnlijk de meest voorkomende factor gevoelig voor metabole ziekten veroorzaken. Vele studies hebben gemeld directe verbindingen tussen voeding en de structuur van de darm microbiome in muismodellen. Een recent voorbeeld opgemerkt dat microbioom structuur snel verschuift naar aanleiding van een verandering van een laag-vet, plantaardig dieet met een hoog-suiker, hoog-vet dieet, het wijzigen zowel de beschikbare metabole routes en actuele genexpressie [63, 64] . Bovendien is de verandering van eetgewoonten geassocieerd met een verhoogd darmpermeabiliteit aan LPS leidt tot een toestand van insulineresistentie [3, 65]. Deze laatste studie verder aangetoond dat de verandering in darmflora werd ook geassocieerd met een andere gevoeligheid voor antibiotica [65]. De hoog-vet dieet-gevoede muizen waren gevoeliger voor de behandeling met antibiotica wat suggereert dat de nieuwe microbiële ecologie dat de dieetbehandeling volgt was zeer broos. In de afgelopen decennia is het aandeel van vet in de voeding grotendeels vervangen die van voedingsvezels, waardoor dus ook de prebiotische effect van de wijze waarop deze twee. Daarom heeft darmflora, althans gedeeltelijk, veranderd in reactie op de nieuwe voeding gewoonte. Kunnen dus nieuwe antigene determinanten en risicofactoren voor immunomodulatie en het optreden van diabetes worden gerelateerd aan de verandering in microbiome. Dit sterk beïnvloed de structuur van de microbiota in een enkele dag, veranderde de vertegenwoordiging van metabole routes in de microbioom, en veranderde microbioom genexpressie. Gehumaniseerd muizen gevoed met het westerse dieet is toegenomen adipositas. Deze eigenschap is overdraagbaar via microbiota transplantatie. Opgemerkt zij dat in humane studies waar de meeste analyses gemaakt van fecale flora, de resultaten vertegenwoordigen waarschijnlijk 5-20% van de totale darmflora. Daarom talrijke bacteriesoorten moeten nog worden beschreven.
De behandeling van metabole ziekten is een uitdaging. Bariatrische chirurgie is momenteel de enige beschikbare behandeling voor morbide obesitas die consequent bereikt en in stand houdt substantieel gewichtsverlies [8, 53]. Het wordt steeds een veel gebruikt procedure, zelfs voor diabetes en obesitas patiënten met matige overgewicht. Verschillende chirurgische procedures zijn ontwikkeld in de afgelopen 50-60 jaar. De Roux-en-Y gastric bypass (RYGB) omvat het creëren van een kleine (ongeveer 15-30 ml) pouch maag van de bodem van de maag, vandaar de distale en proximale maag dunne darm zijn omzeild door aan het distale uiteinde van de mid -jejunum aan de proximale maag zakje (het creëren van de Roux ledematen) en vervolgens opnieuw bevestigen van de gal en pancreas ledematen op een specifieke locatie langs de Roux ledematen [53, 66]. Deze zeer efficiënte procedure kan de remissie van diabetes en vervolgens een belangrijke vermindering van het lichaamsgewicht. Een belangrijk gevolg is de snelle verandering in enteroendocrine functies, zoals de uitscheiding van darmhormonen zoals GLP-1, waarvan de oorsprong is onbekend [67]. Het is echter ook geassocieerd met een drastische verandering van microbiota, waardoor de oorzaak van de entero-endocriene verandering [68, 69]. Met name de firmicutes waren dominant in een normaal gewicht en met obesitas individuen, maar aanzienlijk gedaald in het post-gastric-bypass individuen, die een evenredige toename van Gammaproteobacteria. Interessant is dat het aantal H2-producerende familie Prevotellaceae sterk verrijkt in obesitas en de Archaea leden van de orde Methanobacteriales, die H2-oxiderende bacteriën methaan. De rol is onbekend maar kunnen worden gekoppeld aan een mechanisme belangrijk om de energieopname door de menselijke dikke darm in obese personen. Interessant is dat obese diabetische patiënten en ook gekenmerkt door een vermindering van de anti-inflammatoire bacterie F. prausnitzii [68] in vergelijking met patiënten met overgewicht. Sommige moleculen die met ingang van intestinale microbiota werden gekenmerkt met behulp van NMR-gebaseerde metaboloom analyses [70]. Darmflora afgeleide metabolieten zoals hippuurzuur, Trigonelline, 2-hydroxyisobutyraat, en xanthine meest heeft bijgedragen aan de classificatie-model en waren verantwoordelijk voor de discriminatie tussen obesitas en mager individuen. Bovendien is de typische obesitas metabotype (samentrekking voor metabole fenotype) verloren na gewichtsverlies veroorzaakt door bariatrische chirurgie. De causale rol van deze moleculen niet aangetoond worden maar kan tenminste gesuggereerd dat zij biomarkers vertegenwoordigen.
Mechanismen waardoor darmflora kan leiden tot de ontwikkeling van hyperglycemia en vetopslag
De opslag hypothese
Intestinale microben gebruiken voedingsstoffen en produceren metabolieten die een breed spectrum van menselijke metabole fenotypen, waaronder gevoeligheid voor aandoeningen zoals obesitas [3, 42, 49, 71], insulineresistentie [3], metabool syndroom [89], leversteatose [72 beïnvloeden , 73]. Met behulp van metagenomic en biochemische analyses, Gordon’s groep lieten zien dat deze veranderingen de metabole potentieel van de darmflora van obese muizen [43] beïnvloeden. De “zwaarlijvige microbioom” heeft een verhoogde capaciteit om energie te oogsten uit de voeding [49]. Dit werd echter mechanisme bestreden door andere studies waaruit blijkt dat de verhouding tussen de microbiële samenstelling en energie vangstcapaciteit is complexer dan vroeger beschouwd [74]. Een groot punt was dat deze eigenschap is omdat overdraagbare kolonisatie van kiemvrije muizen met de microbiota van obese muizen resulteerde in een significant grotere stijging van de totale lichaamsvet (tot 40%) dan met de kolonisatie microbiota van een lean muis [ 49, 71]. Derhalve, in tegenstelling tot muizen met een darmflora worden kiemvrije dieren beschermd tegen obesitas die zich na het nuttigen van een Westerse stijl, vetrijke, suiker-dieet [71]. De aanhoudende lean fenotype is geassocieerd met verhoogde skeletspieren en de lever niveaus van gefosforyleerd AMP-geactiveerd proteïne kinase (AMPK) en downstream betrokken in vetzuuroxidatie zoals acetylCoA carboxylase en-carnitine palmitoyltransferase. Verder is de kiemvrije muizen verhoogde expressie van de intestinale vasten geïnduceerde adipocyte factor (FIAF) de peroxisomal proliferator-activated receptor gamma coactivator-1alpha (PGC-1alpha) induceert. Zo, de auteurs gesuggereerd dat kiemvrije muizen werden beschermd tegen dieet-geïnduceerde obesitas door twee onafhankelijke, maar complementaire mechanismen die leiden tot een verhoogde stofwisseling van vetzuren.
De inflammatoire hypothese
Metabole ziekten worden gekenmerkt door een low-grade ontsteking en de rol van het aangeboren en adaptieve immuunsysteem van groot belang [37, 38, 62, 75-78]. De oorsprong van de factoren ontsteking vóór het begin van obesitas of diabetes blijft onbekend. Eerst voorgesteld de lipopolysacchariden (LPS) die zeer inflammatogenic component van de celwand van Gram-negatieve bacteriën causaal betrokken bij het begin van de laaggradige ontsteking in reactie op een vet verrijkt dieet [3]. Bacteriële fragmenten worden herkend door Toll-like receptoren (TLR’s) die een geconserveerde familie van integraal membraan patroonherkenning receptoren die een cruciale rol spelen in het aangeboren immuunsysteem, dat is het begin van de afweer tegen binnendringende ziekteverwekkers hebben, maar zijn ook nodig voor intestinale homeostase [79]. Andere intracellulaire receptoren, zoals de nucleotide-bindende domein Oligomerisatie (NOD)-like receptoren (NLRs) van bacteriële DNA of NOD1 en NOD2, andere fragmenten zoals peptidoglycanen, betrokken bij aangeboren immuniteit [80, 81] en geacht als doel om ontsteking te controleren. Muizen een vetrijk dieet voor een korte periode van 2 weken waar gekenmerkt door een gematigd 2-3-voudige toename in bloed LPS gedefinieerd als metabole endotoxemia [3]. Het blijft binnen dit bereik, dat wil zeggen ver van wat waargenomen tijdens een acute infectie. Derhalve op basis van deze constante hoge concentratie van plasma LPS, vetweefsel, lever en spierontsteking ontwikkeld [3, 82]. De causaliteit van deze bacteriële factor werd aangetoond, aangezien een lage snelheid continu infusie van LPS geïnduceerde meeste vroege factoren recapituleren metabool syndroom en omgekeerd muizen verwijderd uit de LPS receptor TLR4 of deel van TLR4 machines zoals CD14, weerstand het optreden van de ziekte [83]. Bovendien adipocyten behandeld met LPS een ontsteking [84]. Tal van cijfers dat de oorspronkelijke plaats van de ontsteking inderdaad het vetweefsel, maar recente studies, met behulp van conventionele en kiemvrije muizen, suggereerde dat het zou kunnen worden gelokaliseerd in de darm, waar HF westers dieet en darmbacteriën samenwerken om darmontsteking, wat bijdraagt bevorderen de progressie van obesitas en insulineresistentie [85]. De rol van darmflora werd aangetoond aangezien een chronische behandeling met antibiotica verminderde de intensiteit van de ziekte in vetrijke dieet en ob / ob muizen [65, 86]. Plasma LPS concentratie kan worden beschouwd als een risicofactor was aanwezig in overmaat in het bloed van patiënten ogenschijnlijk gezonde voeding meer vetten dan koolhydraten of eiwitten [82]. De verhoogde plasma concentratie kan acuut LPS geïnduceerd worden door een absorptie van lipiden in human [87] en in muizen [3] en lijkt op een verhoogde darmpermeabiliteit afhankelijk via een GLP-2 afhankelijk mechanisme [88]. Andere bewijzen toonden aan dat de TLR5 receptor omgekeerd is het beschermen tegen het metabool syndroom, omdat muizen die genetisch deficiënt zijn in TLR5 tentoongesteld vraatzucht en ontwikkeld kenmerk kenmerken van het metabool syndroom [89]. TLR5 helpt in eerste instantie te verdedigen tegen infecties. Daarbij werd aangetoond dat TLR5 controles hyperfagie, hyperlipidemie, hypertensie, insulineresistentie, en verhoogde adipositas de mechanismen die ontsteking. Een ander fenotype was dat de metabole eigenschappen gecorreleerd met veranderingen in de samenstelling van de darmflora. Het fenotype overdraagbaar was sinds de transplantatie van de microbiota van TLR5-deficiënte aan kiemvrije muizen resulteerde in obesitas en verminderde insulinegevoeligheid. Deze bevindingen tonen aan dat de modulatie van het immuunsysteem ontvangende lichaam beïnvloedt door het veranderen darmflora.
Rekening houdend met wat hierboven gemeld, een belangrijke zaak is om te begrijpen hoe bacteriële fragmenten en LPS de doelorganen en ontsteking te bereiken. LPS moleculen worden meegenomen in het bloed voornamelijk door lipoproteïnen wanneer in de lever zij zijn voorgesteld induceren hepatitis [90]. Bovendien is een antibacteriële behandeling verminderde de ziekten [91]. Daarom is gesuggereerd dat LPS kan worden geabsorbeerd door de darm tijdens de synthese van chylomicrons [92, 93] dan uitgewisseld met andere lipoproteïnen [94] die chronisch kan worden getransporteerd naar doelwit weefsels zoals lever [95] of bloedvaten [96] en ontsteking. Echter, in een acute situatie, kan een flits toediening van lipoproteïnen bufferen plasma LPS en het verminderen van de impact ervan op de acute fase ontsteking [97, 98]. Geen hypothese is tot nu toe beschikbaar is voor andere bacteriële fragmenten.
Al met al, de enorme diversiteit van de intestinale microbiota kunnen meerdere hypothesen over de moleculaire mechanismen die verantwoordelijk zijn voor metabole ziekten. Zeker, ontsteking worden gehouden. Men zou ook kunnen suggereren dat inname van voedsel en energie-opslag moeten worden betrokken bij het beeld.
Gut microbiota en vetstofwisseling
Conclusies uit de vorige paragraaf blijkt dat intestinale microbiota, die sterk van invloed op de opslag van vet in het wit vetweefsel, kan net zo goed strak vetstofwisseling en de gevolgen daarvan te regelen op hart-en vaatziekten. Deze hypothese is gemakkelijk intuïtief omdat de darm is de toegangsdeur van lipide. Microbiota, hoewel aanwezig in lage concentratie in het duodenum en jejunum (honderdvier-honderdenvijf cel / ml), waar de meeste lipiden worden opgenomen, worden de informatie darmcellen met lipide metabolieten. Anders een overmaat lipiden dus niet opgenomen, zou het voeden van de microbiota in de dikke darm op toch informatieve metabolieten ook. Bewijzen toonde aan dat conventionele muizen met normale microbiota gekenmerkt door een verhoogde productie van energie metabolieten, bijvoorbeeld, pyrodruivenzuur, citroenzuur, fumaarzuur, appelzuur en, in vergelijking met kiemvrije muizen [99]. Omgekeerd werden plasmaspiegels van cholesterol en een aantal soorten lipiden in het serum triglyceriden en fosfatidylcholine verminderd met de microbiome terwijl zij verhoogd in het weefsel, zoals het vetweefsel en de lever. Dit suggereerde dat de klaring van lipiden werd verhoogd door de microbiota. De mechanismen blijven onbekend maar waarschijnlijk door de verandering in de bacteriële genera en de overeenkomstige microbiome, al dan niet aanwezig in de flora van het diabetes of obesitas dieren. Zelfs cholesterol in de darmen metabolisme wordt beïnvloed door de darmflora. In kiemvrije ratten, lever microsomale hydroxylering van steroïde hormonen en lithocholinezuur is efficiënter dan de conventionele tegenhangers [100]. Een precieze voorbeeld is de ontdekking van een Bacteroidetes D8, een cholesterolverlagend bacterie van humane intestinale oorsprong, die werd geïsoleerd uit een senior mannelijke vrijwilligers met een hoge capaciteit om luminale cholesterol te coprostanol [101]. Tientallen jaren geleden werd de rol van darmflora betreffende de controle van het vetmetabolisme indirect gesuggereerd omdat werd aangetoond dat galzuren kunnen worden gemetaboliseerd door de flora [102]. In kiemvrije ratten, de hoeveelheid urobiline en stercobilin vrijwel verwaarloosbaar waaruit blijkt dat bilirubine wordt gereduceerd tot urobilins de darmflora uitsluitend. De kolonisatie met verschillende bacteriestammen gebleken dat het metabolisme van galzuren anders werd beïnvloed door het type darmflora. In alle gevallen was monocolonization niet volledig kunnen weer een normaal galzuur metabolisering.
Verrassenderwijs heeft het effect van darmflora op systemisch metabolisme (Fig. (Fig.1) 1) aangetoond zelfs met betrekking tot lipide homeostase in niet-metabolisch actieve organen zoals het oog [103]. In een recente studie, Oresic et al.. vergeleek de lipide structuur van ogen afgegeven vanaf kiemvrije en conventionele muizen. De auteurs analyseerden zowel lens en netvlies lipidome door massaspectrometrie (MS) uitgevoerd in ion-negatieve modus (ESI-) en in totaal 140 en 276 lipiden waren respectievelijk gedetecteerd. De belangrijkste bevinding was de microbiota aangedreven daarbij de algemene fosfatidylcholinen, die een verhoogde blootstelling voorgesteld oxidatieve stress in conventionele muizen in vergelijking met kiemvrije muizen.
Fig. 1
Multiple-sited impact van darmflora op tal metabolisme. Gut microben is aangetoond of voorgesteld om een impact hebben op het vetweefsel en de lever vet opslag, skeletspier energiemetabolisme, vet metabolisme in de lever en hepatische steatose, atherosclerose hebben …
Echter, in de zoektocht van mechanismen die darmflora, vetstofwisseling en vaatziekten recente vinden tonen een sterke moleculaire hypothesen [104]. Voor, het bespreken van deze onderwerpen, is het opmerkelijk dat hart-en vaatziekten zijn gekoppeld aan infectie tientallen jaren, door het vergroten pro-atherosclerotische veranderingen in de vasculaire cellen [105]. Een Microbiome gevonden in atherosclerotische plaques omdat bacterieel DNA kunnen worden geïdentificeerd in meer dan 50% van alle plaques [106] en de oorsprong kan intestinale of oraal [107]. De vasculaire risicofactoren inderdaad verhoogd populatie studies waarin de plasmaconcentratie van LPS werd verhoogd (Fig. 2) [104, 108, 109]. Omgekeerd anti-LPS moleculen zoals oplosbare CD14 tegen stijfheid van de aorta en dus een verminderde vaatfunctie [110]. Daarom kunnen antibiotica therapieën hebben een positief effect op de vasculaire functie [111]. De moleculaire controle van de productie van atherosclerotische plaques door factoren van intestinale origineel is onlangs in een studie waarin de auteurs gebruikten een metabolomics benadering onpartijdige kleine moleculen metabolische profielen in plasma CVD risico voorspellen genereren. Drie metabolieten van de voeding lipide fosfatidylcholine-choline, trimethylamine N-oxide (TMAO) en betaïne werden geïdentificeerd en vervolgens naar risico voor CVD voorspellen onafhankelijke grote klinische cohort. Aanvulling van de voeding en kiemvrije muizen studies bevestigden een cruciale rol voor de voedings-choline en darmflora in TMAO productie. Het was gekoppeld aan cholesterol accumulatie en vorming van schuimcellen macrofaag. Vandaar dat de rol van monocyten / macrofagen is belangrijk in low-grade ontstekingsziekten, omdat een toename van het aantal gunsten coronaire collaterale groei bij type 2 diabetes patiënten [112]. Uiteindelijk de onderdrukking van intestinale microflora in atherosclerose-gevoelige muizen remde dieet-choline-enhanced atherosclerose. Daarom wordt microbiota van de darm of orale oorsprong nu zeker erkend als een risico en een oorzakelijke factor van de cascade van gebeurtenissen die leiden tot atherosclerose. Een interessante hypothese zou zijn dat de microbioom kon gastheer genexpressie bedienen via miRNA [113]. Vergelijkende profilering van miRNA expressie met behulp van miRNA arrays van conventionele en kiemvrije muizen bleek dat mmu-mir-665, die werd ontregeld tijdens kolonisatie, Abcc3 expressie down-gereguleerd door direct gericht op de Abcc3 3′-UTR [113]. De rol van miRNA op endotheliale metabolisme is elders aangetoond [114] en daarom kon men suggereren dat darmflora kan endotheelfunctie en menselijke atherosclerotische laesies [114] regelen. Echter, de algemene controle van de ontsteking gemedieerd door het aangeboren en adaptieve immuunsysteem zeker regelen de agressiviteit van de darmflora. Bijgevolg zal de waarheid steunen op een set markers geassocieerd darmflora als risicofactor, de regulerende rol van het immuunsysteem en de genetische achtergrond van het individu.
Fig. 2
De inflammatoire branden: darmflora dysbiose en de oorsprong van metabole stoornissen. De oorsprong van metabole ziekten is multifactorieel, maar de impact van schadelijke voedingsgewoonten is zeker de belangrijkste factor die verantwoordelijk is. Dit wijzigt direct …
Gut microbiota als een regulator van leversteatose
Hepatische steatose is een van de belangrijkste complicaties van abdominale obesitas, insulineresistentie en type 2 diabetes [115 tot 117]. Ongeveer 60-80% van deze patiënten een stadium van de ziekte van 1 tot 5 waarbij de laagste limiet komt overeen met de ophoping van lipiden en vervolgens tot ontsteking, apoptose, fibrose, leverfalen of kanker. Het wordt nu steeds een prioriteit voor Europa door de lancering van de Europese programma’s voor onderzoek te financieren. Het argument dat intestinale microbiota betrokken zou zijn bij de activering van leversteatose is afkomstig van waarnemingen die conventionalized dieren 40% meer lichaamsvet dan kiemvrije dieren [42] hebben. Verschillende moleculaire mechanismen zijn voorgesteld. Een verhoogde intestinale productie van korte keten vetzuren worden die meer energie naar de lever [49, 71]. De genetische achtergrond geeft een gevoeligheid voor het ontwikkelen van leversteatose zoals in de SV129S6 muis [72]. In dit geval zijn de multivariate statistische modellen van de metaboloom spectra van urine aangetoond dat de genetische aanleg van de muizenstam om leversteatose is geassocieerd met storingen van choline metabolisme [72]. Werd aangetoond dat de symbiotische darmflora de choline omgezet in methylaminen leidt tot lage circulerende niveaus van plasma fosfatidylcholine en hoge uitscheiding van methylamines (dimethylamine, trimethylamine, en trimethylamine N-oxide), die de biologische beschikbaarheid van choline vermindert en bootst het effect van choline-deficiënt dieet, waardoor NAFLD [72, 73]. Echter, dit model niet om een spil die de leverziekte wordt geassocieerd met een toestand van ontsteking. De moleculaire inflammatoire mechanisme kan worden toegeschreven aan plasma LPS die toeneemt bij patiënten met levercirrose en in de ader hepatoportal volgende alcoholgebruik [3, 118, 119]. Dit mechanisme was afhankelijk van TLR4 en initieert ontstekingen [120]. Echter, in afwezigheid van alcoholgebruik, plasma-LPS concentratie een matige verhoging in het bloed van muizen die een vette lever ontwikkelen, zoals de hoog-vet dieet-gevoede muizen [3]. Dit proces kenmerkt metabole endotoxemie en wordt beschouwd als een factor triggering van ontsteking en metabolische ziekten. Metabole endotoxemie werd gekoppeld aan een verhoogd darmpermeabiliteit [65]. De behandeling van de muizen een vet dieet verrijkt met antibiotica verminderde de metabole endotoxemie en de ophoping van vet in de lever en de lokale inflammatoire status [65, 86]. Bovendien het inflammatoire mechanisme, is voorgesteld dat darmflora zou vetophoping worden steeds in de lever via een mechanisme waarbij de regulatie van FIAF [42, 71]. Dit lid van de angiopoietine-achtige eiwitfamilie selectief onderdrukt in het darmepitheel van normale muizen door darmflora. FIAF is een circulerende lipoproteïne lipase inhibitor en onderdrukking is essentieel voor micro-geïnduceerde depositie van triglyceriden in adipocyten. Inderdaad, in de obese diabetische ob / ob muis, de overeenkomstige microbiome een verhoogde capaciteit om energie uit de voeding verder aanwakkeren van de lever met koolstofresten gegenereerd door bacteriële fermentatie van onverteerbare vezels. Derhalve is na een verandering van darmflora wordt voorgesteld dat zowel verhoogde voedingsvezels fermentatie en de inductie van metabole endotoxemie leidt tot lipiden ophopen in lever en ontsteking. Deze hypothese zou passen bij de snelle ontwikkeling van leversteatose ondersteunend de verandering in voedingsgewoonten en dus van de intestinale microbiota.
Gut microbiota en type 1 diabetes
Type 1 diabetes is een auto-immuunziekte die wordt veroorzaakt door de specifieke vernietiging van de insuline uitscheidende endocriene pancreatische beta cellen door T-lymfocyten [121]. Bijgevolg is een progressieve maar absolute insulinopenic toestand treedt op in de volgende maanden en jaren. Dit mechanisme is onomkeerbaar, maar kan worden voorkomen door immunosuppressiva bij gebruik vroeg genoeg [122, 123]. De antigenen waarop het immuunsysteem gedeeltelijk zijn beschreven en hebben betrekking op de erkenning van glutaminezuurdecarboxylase (GAD) -64 eiwitten. T-lymfocyten dringen de eilandjes van Langerhans, zoals beschreven door een insulitis, en steeds meer vernietigen de insuline afscheidende cellen alleen [121]. Wat blijft onbekend is het proces dat voorkomt de vernietiging van de juiste autoreacting T-lymfocyten of het systeem dat de auto-immuunreactie door miskenning van de zelf-antigenen zoals GAD64 toelaat. Men zou kunnen suggereren dat de rijping van T-lymfocyten zou worden aangetast. Recente gegevens toonden de cruciale rol van de gastro-intestinale microbiota in de bescherming of het activeren van type 1 diabetes [57, 124]. De diermodellen geschikt voor dergelijke studies zijn de niet-obese diabetische (NOD) muis stam, of de BB rat. In beide modellen worden de doel-pancreas insuline producerende beta-cellen aangevallen en verwoest door geactiveerde immuuncellen, wat leidt tot type-1 diabetes. De ontdekking van de rol van darmflora afkomstig van de hygiënische hypothese [23] na de waarneming dat de incidentie van spontane T1D in de NOD muizenkolonie kan worden beïnvloed door de microflora in de stalruimte inrichting [125] of door blootstelling aan microbiële stimuli zoals injectie met Mycobacterium of verschillende microbiële producten [126, 127]. In de menselijke, is de incidentie van type 1 diabetes toegenomen in de afgelopen decennia in de ontwikkelde landen waar de milieu-omstandigheden drastisch zijn veranderd [honderdachtëntwintig-dertienhonderdeneen]. Deze hypothese suggereert dat bacteriële antigenen zou worden gepresenteerd door het aangeboren immuunsysteem op de T-lymfocyten heel vroeg in het leven ondersteunen het idee dat immunostimulatie kan de rijping van de postnatale immuunsysteem [132, 133] ten goede komen. Derhalve bij miskenning van het bacterieel antigeen, wordt het adaptieve immuunsysteem verergert de agressiviteit tegen de pancreatische cellen en vernietigen. Bijgevolg zou de erkenning van bacteriële determinanten van intestinale microbiota een uitlokkende factor van auto-immuunziekte. De Toll-like receptoren (TLR’s) zijn aangeboren patroonherkenning receptoren die betrokken zijn bij de afweer die de controle over commensale bacteriën en weefsel integriteit [79, 134] te houden. De overeenkomstige signalerend molecuul is MyD88 adapter derhalve muizen die deze moleculaire mechanismen tegen insulitis [57]. Dit is afhankelijk van de commensale bacteriën omdat kiemvrije MyD88 knockout muizen ontwikkelen robuuste diabetes.
Het type darmflora is belangrijk. Die van NOD muizen waarschijnlijk induceren diabetes sinds de kolonisatie van deze muizen met een microbiota van niet-type 1 diabetische muizen verhinderd de incidentie van diabetes in kiemvrije NOD muizen. Derhalve zijn zowel de kwaliteit van de darmflora en de activering van het aangeboren immuunsysteem de controle van de agressiviteit van T-lymfocyten en daarmee de ontwikkeling van auto-immuunziekten. In de BioBreeding rat model van type 1 diabetes, is gebleken dat de Lactobacillus soorten in feces werden negatief gecorreleerd met type 1 diabetes ontwikkeling [135]. Juist, twee soorten de L. johnsonii en L. ruteri verhinderd het effect op diabetes type 1 ontwikkeling [135]. Gesuggereerd werd dat twee stammen van bacteriën veranderingen in de intestinale mucosale eiwit en oxidatieve stress leidend tot lage IFNy geïnduceerd. Bijgevolg waarnemingen na toediening van antibiotica in type 1 diabetes BioBreeding ratmodellen gebleken dat het ontstaan van de ziekte is verminderd, dat de hypothese dat een specifieke darmflora kan autoimmuniteit induceren versterkt [136].
In totaal techniek darmflora door het gemiddelde van prebiotica, probiotica, en voedsel complement, of door bacteriële-afgeleide immunotherapeutische strategieën kunnen beneficiate de preventie van type 1 diabetes (afb. (Fig.33).
Fig. 3
Therapie strategieën uitdagend darm microben. De ontdekking van de rol van darmflora betreffende de controle van metabole ziekten opent vele therapeutische strategieën zoals prebiotica, probiotica en immuunmodulatie. Ook kan de generatie …
Gut microbiota en parodontitis
Cross-sectionele studies suggereren een sterke associatie tussen systemische ziekten en orale infectie, zoals parodontitis [137 tot 140], die wordt beschouwd als de zesde complicatie van diabetes mellitus [141, 142]. Parodontitis is een veel voorkomende chronische multifactoriële infectie wordt gekenmerkt door een ontstekingsreactie tegen een specifieke microbiota mond. Deze stelt een complex biofilm die vooral van Gram-negatieve bacteriën samengesteld in de subgingivale micro [143, 144]. Op basis van dit aspect, Saito et al.. [145] verondersteld dat de specifieke correlatie tussen de periopathogens en de ontwikkeling van obesitas ook kan worden geassocieerd met veranderingen in plasma LPS concentratie. Kan vooral de endotoxine uit de Gram-negatieve pathogenen verantwoordelijk op afstand effecten van parodontitis op gezondheidsproblemen van [145]. LPS release in de systemische circulatie is direct van invloed organen zoals de lever, de longen, vetweefsel, skeletspieren en het hart [146]. De uitdaging van deze bacterieel antigeen op meerdere plaatsen orgaan kan een ontstekingsreactie veroorzaken, resulterend in insulineresistentie, wat uiteindelijk leidt tot metabole stoornissen [38]. Met betrekking tot andere chronische ziekte geassocieerd met een low-grade ontsteking zoals atherosclerose, recente gegevens laten zien dat bacteriën uit de mondholte kan samenhangen met ziekte markers van vaatziekten [107]. Door zich te richten de 16S rRNA V1-V2 regio door pyrosequencing en door het toepassen van qPCR analyse, data geïdentificeerd gedeeld sequenties die behoren tot de Veillonella en Streptococcus groepen van sequenties tussen orale en atherosclerotische plaque monsters binnen dezelfde personen. Bovendien, in deze studie principal component analyses suggereerden dat de overvloed aan Fusobacterium, een van de eerste kolonisten in periodontale plaque, kan direct de niveaus van totaal cholesterol en LDL beïnvloeden. Dit bewijs toont de translocatie van orale bacteriële naar de systemische circulatie. Dus in het licht van de bovenvermelde resultaten hebben vele studies aangegeven dat de behandeling parodontitis doordat orale bacteriële pathogenen [147-149] kan metabole controle bij diabetische patiënten.
Het gebruik van Pre-, Pro-, en synbiotische te voorkomen of te behandelen metabolische problemen
Tegenover de epidemie van obesitas en diabetes, moet men erkennen dat preventieve strategieën moeten worden toegepast. Tot nu toe zijn juiste voedingsgewoonten alleen nauwelijks toegepast door personen terwijl meer en meer tieners zijn onder de invloed van vet-en suiker-verrijkte voeding. Dus voor dit maatschappelijke probleem nieuwe strategieën worden voorzien.
Een manier zou zijn om onze darmflora te herstellen door het gemiddelde van pre-of pro-biotica. Wat prebiotische er nu veel belangstelling manipulatie van de microbiota samenstelling om het mogelijk gunstige aspecten te verbeteren. De prebiotische aanpak dicteert dat niet-levensvatbare ingrediënten worden speciaal gefermenteerd in de dikke darm door de inheemse bacteriën gedacht te worden van positieve waarde, bijvoorbeeld bifidobacteriën, lactobacillen [150, 151]. Elk ingrediënt dat in de dikke darm komt is een kandidaat-prebiotisch [152]. De meeste huidige aandacht en succes zijn afgeleid met behulp van niet-verteerbare oligosachariden uit fructose, xylose, soja, galactose, glucose, mannose en met verschillende werkzaamheid op stofwisselingsziekten. Het werkingsmechanisme blijft onduidelijk, maar kan worden gerelateerd aan de regulering van intestinale mucosale biologie, waar de intestinale mucosa werd gekenmerkt door een hoger villi, diepere crypten, verhoogd aantal slijmbekercellen en een dikkere slijmlaag op colonepitheel [153]. De remming van intestinale permeabiliteit voor middelen zoals LPS is onlangs voorgesteld een formule die kan worden waarbij de afscheiding van enterische peptide zoals glucagon-achtig peptide 2 [88]. Uiteindelijk konden intestinale entero-endocriene functies ook worden gericht op prebiotica [154]. Fructoligosaccharides verhogen de productie van glucagon-achtige peptide 1 dat insulinesecretie kunnen bevorderen en op darm-hersenen as voor de regulatie van glucose metabolisme [155].
Andere strategie is het gebruik van bacteriën, dat wil zeggen probiotische een gezonde darmflora herstellen. Verschillende geslachten worden momenteel gebruikt en onder hen de meest voorkomende zijn bifidobacteriën en lactobacillen met verschillende voordelen voor de gezondheid. De geïdentificeerde werkingsmechanismen zijn tal van [156]. Ze lijken ten minste gedeeltelijk verband met de wijziging van de hechting van bepaalde bacteriestammen de mucosa [157], de invloed van het enterale immuunsysteem door de productie van IgA [158] of de inductie van anti-inflammatoire moleculen als Il10 [honderdnegenenvijftig-honderdeenenzestig], of de regeling van de darmpermeabiliteit [162]. Een recent bewijs heeft duidelijk aangetoond dat een specifieke stam, namelijk de B. longum kan ontsteking van de darm te verminderen door het produceren grote hoeveelheid acetaat uit de fermentatie van voedingsvezels [163]. Acetaat interageert met het enterale immuunsysteem om de synthese van regulerende T-lymfocyten bevorderen. Andere mechanismen kunnen het produceren van single-bacteriële moleculen zoals polysaccharide A van Bacteroides fragilis die moet pro-inflammatoire interleukine-17 te onderdrukken door intestinale immuuncellen [164]. De metabole gevolgen van het gebruik van probiotica zijn talrijk. De meesten van hen betrekking hebben op de vermindering van het lichaamsgewicht [165], niet-alcoholische leververvetting [zestienhonderdenzes-honderdachtenzestig], of glycemie en insuline resistentie [169, 170].
Om het licht van de gunstige effecten van pre-en pro-biotica, is een nieuwe strategie combinatie van beide werkwijzen nu omhoog en gedefinieerd als “synbiotische.” Inzicht in de mutualistische verbinding tussen darmflora en gastheer fysiologie voor de controle van de stofwisseling is cruciaal in de zoektocht naar mechanismen die verantwoordelijk zijn van de dramatische verhoging van hart-en vaatziekten [2, 3]. In dit scenario kort een synbiotische strategie bleek geschikt voor het verbeteren van het lipidenprofiel van hypercholesterolemische mannen en vrouwen. In deze studie werden patiënten kregen een combinatie van Lactobacillus acidophilus CHO-220 en inuline [150, 151] of placebo gedurende 12 weken. Ondanks het gebrek aan effect op het lichaamsgewicht en de energie-inname, de synbiotische behandeling verminderde zowel de totale en LDL-cholesterolgehalte in het plasma via een mechanisme waarbij lipide transporters [39]. Aangezien triglyceriden concentratie in lipoproteïnen wordt beschouwd als belangrijkste risicofactoren voor atherosclerose, de auteurs gesuggereerd atheropreventive de rol van synbiotische strategie.
Conclusies: therapeutische en het voorkomen van de huidige en toekomstige mogelijkheden
We worden nu geconfronteerd met een nieuw tijdperk waarin we zullen moeten om de rol van een nieuw orgel rijk aan meer dan 3 miljoen genen te begrijpen: darmflora. Het is waarschijnlijk dat deze metagenoom zal de basis van vele nieuwe therapeutische benaderingen ter behandeling en voorkoming stofwisselingsziekten en de overeenkomstige cardiovasculaire gevolgen. Door het begrijpen van deze nieuwe ecologie strategieën op basis van prebiotica, probiotica kan, zelfs gerichte antibiotica worden overwogen. De identificatie van de eukaryote genen gereguleerd door darmflora zal ook worden beschouwd als nieuwe doelwitten waartegen de farmaceutische bedrijven moeten in staat zijn om het ontwerp verbindingen. Uiteindelijk wordt darmflora beschouwd als een groot aantal antigenen en sommige kunnen dienen als basis immunotherapeutical strategieën te voorkomen of te behandelen. Diagnostische kits aan patiënten identificeren die risico lopen of te classificeren en stratificeren de ziekten zal nuttig zijn voor de arts beter aan te passen van de therapeutische strategie.
Ga naar:
Open Access
Dit artikel wordt gedistribueerd onder de voorwaarden van de Creative Commons Naamsvermelding-Niet Commercieel License waarop een niet-commercieel gebruik, distributie en reproductie in elk medium maakt, op voorwaarde dat de oorspronkelijke auteur (s) en de bron worden gecrediteerd.
Waarom ONDERZOEK naar de oorzaak van DIABETES, terwijl wij kPNI-ers allang weten wat de oorzaak is. Dat bedoel ik nu GELDVERKWISTING.
Dit schreef ik eerder deze week “Insulineresistentie” wat is dat….. IK HERHAAL het komt hier op neer; omdat wij Nederlanders teveel en tevaak koolhydrat eneten, (in Limburg merk ik in mijn praktijk “Aurora” dat de limburgers veelongezonder leven dan de rest van het Land, wat ook in de krant stond vanochtend) wordt een receptor op de cel mindergevoelig voor de insuline. (taak van insuline is om glucose naar de cel te transporteren) Glucose wordt door een stofje IN de cel getransporteerd…en dmv de citroenzuurcyclus….energie uitgehaald.Maar omdat dit opgegeven moment veel minder gebeurd wordt er minder energie naar de cel getransporteerd. Wordt glucose door de lever omgezet in glycogeen (glucose moet toch verwerkt worden–>opslag als glycogeen in de spieren en lever) en als dat verzadigt is in vetzuren VETTEN.”SUIKERZIEKTE de cel is helemaal ongevoelig geworden voor INSULINE–> SUIKERZIEKTE.