Probiotica is door de World Health Organization gedefinieerd als “levende kleine organismen die een positieve uitwerking hebben op de gezondheid van de mens wanneer deze in grote hoeveelheden worden ingenomen”. Probiotica zijn dus zogenaamde “goede bacteriën” en de bacteriën die worden gebruikt in probiotica zijn meestal gelijk aan de bacteriën die normaal gehuisvest zijn in de darmen. Deze goede bacteriën doen wat extra’s voor het lichaam en dragen bij aan een goede darmflora. De term darmflora wordt gebruikt om de miljarden bacteriën aan te duiden die in de darmen leven. Een goede darmflora is onder andere belangrijk voor de spijsvertering welke wordt ondersteund door alle bacteriën die in de darmen zitten. Een evenwichtige darmflora is eveneens belangrijk voor een goed werkend immuunsysteem en goede stoelgang.
Prébiotica
Prébiotica (inuline en fructo-oligosacchariden (FOS)) zijn onverteerbare voedingsvezels en zorgen er onder andere voor dat lactobacillen en bifidobacteriën (de goede bacteriën) beter kunnen groeien. Bovenal is uit onderzoek gebleken dat een prebiotica gecombineerd met een probiotica effectiever in het darmkanaal werkt dan de één of ander apart ingenomen.
Ontwikkeling van de darmflora
Tijdens de bevalling wordt de basis voor de darmflora al gelegd. De baby komt voor het eerst met bacteriën in aanraking door contact met het geboortekanaal, de moeder en de omgeving direct na de geboorte. De darmflora wordt voor het grootste deel samengesteld de periode direct na de geboorte. Een kind dat op de reguliere manier ter wereld komt heeft daardoor een andere darmflora samenstelling dan een kind dat via een keizersnede wordt geboren. Na ongeveer 30 dagen is de darmflora van een kind dat middels een keizersnede ter wereld wordt gebracht pas gelijk aan die van een kind die via de reguliere route ter wereld is gekomen. De bacteriën passeren de maag, omdat een baby nog niet veel maagzuur produceert, en vestigen zich op de darmwand. Kolonisatie van de bacteriën kan geschieden doordat er in eerste instantie bacteriën in de darmen aanwezig zijn die al het zuurstof gebruiken. Dit baant een weg voor de bacteriën die kunnen overleven zonder zuurstof (lactobacillen, bifidobacteriën). Vanaf dat moment kunnen deze in aantal toenemen en kan de darmflora zich ontwikkelen met allerlei bacteriën die voor ieder individu anders zijn.
Variatie darmflora
Iedere darmflora is uniek en vergelijkbaar met een vingerafdruk. Doordat de samenstelling zo uniek is, is het lastig te definiëren wat een normale darmflora is. Onderzoek heeft uitgewezen dat ieder individu dezelfde darmflora-kern heeft. Echter beslaat deze kern maar 20% van alle bacteriën die aanwezig zijn in de darmen. De overige 80% is bij een ieder verschillend. Er wordt gesuggereerd dat er maar liefst 1500 verschillende bacteriën aanwezig zijn. Op dit moment wordt daar nog uitvoerig onderzoek naar gedaan.
Bacteriële dysbiose
Om gezond te blijven is het belangrijk de darmflora in balans te houden. Door enkele factoren kan de darmflora echter uit balans raken. Hierbij moet gedacht worden aan het gebruik van antibiotica. Antibiotica staat er om bekend schadelijke bacteriën te doden, maar daarnaast doden ze ook de goede bacteriën in ons maag-darmkanaal. Leeftijd en voeding kunnen eveneens een rol spelen bij de verstoring van de harmonie in de darmen. Bij ouderen bestaat nog maar 1% van de totale darmflora uit bifidobacteriën waar dit bij volwassenen 3-6% is.
Probiotica is belangrijk bij het bestrijden van allerlei verschillende spijsverteringsproblemen. Een goede darmflora is namelijk onmisbaar bij het verteren van ons voedsel. Onze darmflora en enzymen zijn volledig verantwoordelijk voor het verteren van voedsel en opname van voedingsstoffen uit dit voedsel. Wanneer er een bacteriële dysbiose ontstaat en dit systeem niet meer naar behoren functioneert blijven er onnodige hoeveelheden etensresten achter in de darmen, die vervolgens tot ontstekingen kunnen leiden. Hiermee kan probiotica preventief werken en een effectief wapen zijn tegen het ontstaan van ontstekingen en het ontstaan van een verhoogde permeabiliteit van de darmwand met alle gevolgen van dien.
Stammen Probiotica
Het is belangrijk dat de bacteriën in de Probiotica levend aankomen op de eindbestemming, de darmen. Daarom is het belangrijk dat ze zuur-en gal resistent zijn. Hier is uitvoerig onderzoek naar verricht. In tabel 1 is te zien dat vrijwel alle bacteriën zuur-en gal resistent blijken te zijn met Lactobacillus Acidophilus als echte uitblinker op dit gebied.
Tabel 1: In vitro % herstel levende cellen na incubatie in medium met 1 van de factoren
Probiotica hebben de kracht om bepaalde ziekteverwekkers te remmen. Ook in onze stammen is dit nader bekeken bij bepaalde bekende ziekteverwekkers. Uit de resultaten komt naar voren dat vrijwel alle stammen de mogelijkheid hebben de verschillende onderzochte ziekteverwekkers maximaal te remmen. Hoe hoger het cijfer, hoe hoger de remming van die bepaalde ziekteverwekker (figuur 1 & tabel 2).
Figuur 1: De ziekteverwekkers werden gekweekt in een petrischaaltje met Agar-Agar (groeimedium die gebruikt wordt om micro organismen in te kweken). De probiotische stam werd aangebracht in het kleine holletje. In de Controle situatie is er geen remming van de ziekteverwekker (0). In het voorbeeld van goede remming wordt een maximale remming gemeten (3)
Figuur 1: De ziekteverwekkers werden gekweekt in een petrischaaltje met Agar-Agar (groeimedium die gebruikt wordt om micro organismen in te kweken). De probiotische stam werd aangebracht in het kleine holletje. In de Controle situatie is er geen remming van de ziekteverwekker (0). In het voorbeeld van goede remming wordt een maximale remming gemeten (3)
Tabel 2: 0= geen remming van de ziekteverwekker, 1= 8-11 mm, 2 = 11-14 mm 3= >14 mm maximale remming van de ziekteverwekker
T-cel differentiatie
Probiotica kunnen het immuunsysteem activeren door signalen vanuit het lumen via dendritische cellen aan het systeem door te geven. Het immuunsysteem wordt gereguleerd door twee hoofdcategorieën cytokines met verschillende functies en geproduceerd door subsets van T-helper (Th) lymfocyten.
Th1-cellen produceren voornamelijk IL-2 en interferon-γ en reguleren de immuunrespons tegen intracellulaire infecties. Th2-cellen produceren voornamelijk IL-4, IL-5 en IL-13 en zijn juist mediatoren in extracellulaire infecties en atopische ziektes. Regulatorische T-cellen (Treg) zijn de belangrijkste spelers die ervoor kunnen zorgen dat de balans van Th1/Th2 cellen terug naar homeostase gaan. Zoals in de onderstaande afbeelding wordt getoond, activeren de dendritische cellen bepaalde naïeve T-cellen (zijn nog niet in aanraking geweest met een antigeen) die zich vervolgens ontwikkelen tot een van deze meer specifieke T-cellen, die uiteindelijk de bijbehorende cytokines uitscheiden en een immuun reactie uitlokken (figuur 2).
Figuur 2: Dentrische cellen activeren bepaalde naïeve cellen T-cellen
Onze stammen zijn getest op hun mogelijkheid om het afweersysteem positief te beïnvloeden. Alle stammen laten zien dit te doen. Iedere stam had de mogelijkheid naïeve T-cellen te laten prolifereren in een specifieke T-cel. Dit was bevestigd door het meten van cytokine productie.
Bron: beyondmedicine
Graviola: A Novel Promising Natural-Derived Drug That Inhibits Tumorigenicity and Metastasis of Pancreatic Cancer Cells In Vitro and In Vivo Through Altering Cell Metabolism
Pancreatic tumors are resistant to conventional chemotherapies. The present study was aimed at evaluating the potential of a novel plant-derived product as a therapeutic agent for pancreatic cancer (PC). The effects of an extract from the tropical tree Annona Muricata, commonly known as Graviola, was evaluated for cytotoxicity, cell metabolism, cancer-associated protein/gene expression, tumorigenicity, and metastatic properties of PC cells. Our experiments revealed that Graviola induced necrosis of PC cells by inhibiting cellular metabolism. The expression of molecules related to hypoxia and glycolysis in PC cells (i.e. HIF-1α, NF-κB, GLUT1, GLUT4, HKII, and LDHA) were downregulated in the presence of the extract. In vitro functional assays further confirmed the inhibition of tumorigenic properties of PC cells. Overall, the compounds that are naturally present in a Graviola extract inhibited multiple signaling pathways that regulate metabolism, cell cycle, survival, and metastatic properties in PC cells. Collectively, alterations in these parameters led to a decrease in tumorigenicity and metastasis of orthotopically implanted pancreatic tumors, indicating promising characteristics of the natural product against this lethal disease.
Keywords: Pancreatic cancer, therapy, cancer metabolism, natural product
The overall five-year survival rate for pancreatic cancer (PC) patients was 5.5% for the period of 2001–2007, according to the National Cancer Institute (NCI), a statistic that has not varied significantly for over the last four decades [1]. In 2012, it is estimated that 43,920 new PC cases will be diagnosed and approximately 85% of these (i.e. 37,390) will succumb to the disease [2]. The main reason behind the poor prognosis of PC patients is the insidious and sporadic nature of the disease, which is often presented with no specific early clinical symptoms. By the time of diagnosis PC is already in advanced stages (i.e. III and IV) and is resistant to conventional chemotherapy and radiotherapy [3]. Interestingly, even patients diagnosed with stage I PC that have the option to undergo surgery have a 5-year overall survival of approximately 20%, a clear indication of the general failure of current standard treatments for each stage of PC [4, 5]. What is even more alarming, are the statistics that predict possible 55% increase in the expected number of new PC cases by 2030 [6]. Thus, immediate progress must be made in the prevention, early diagnosis, and systemic treatments against this lethal disease.
Gemcitabine has been the standard line of treatment for PC patients for over a decade and is associated with a median patient survival of 5.4 months [7]. Over all these years, numerous clinical efforts have been devoted to improve PC chemotherapy outcomes, but unfortunately no significant improvements have been reported apart from a clinical trial reported in May of 2011 [8]. This phase III clinical trial reported an improved overall survival of PC patients treated with a four-drug chemotherapy regimen comprising fluorouracil, leucovorin, irinotecan, and oxaliplatin (FOLFIRINOX). Nevertheless, a major disadvantage of this novel treatment was its related toxicity, which was noticeably high when compared to PC patients treated with gemcitabine alone. Therefore, novel, alternative PC therapeutics must not only improve the prognosis of PC patients but also minimize any possible toxicity-related side effects that will interfere with the quality of life of PC patients.
It is well known that an increased consumption of fruits and vegetables is associated with a reduced risk of most cancers, including PC [9]. For this reason, the potential of natural products in PC therapies has been widely investigated [10]. While some of these compounds have undergone clinical testing (i.e. curcumin, genistein) and have demonstrated some activity against PC, the poor bioavailability in patients minimizes their therapeutic efficacy. However, as compared with conventional chemotherapeutic drugs, the major benefit of these therapies is the apparent lack of toxicities to healthy tissues. This attracted our attention to find alternative, natural-derived chemotherapeutic drugs in order to improve the prognosis of PC patients. Traditionally, the leaves from the tropical tree Annona Muricata, also known as Graviola or Soursop, have been used for a wide range of human diseases including inflammatory conditions, rheumatism, neuralgia, diabetes, hypertension, insomnia, cystitis, parasitic infections, and cancer [11]. The major bioactive components that have been extracted from different parts of the plant are known as Annonaceous acetogenins. These are derivatives of long chain (C35 or C37) fatty acids derived from the polyketide pathway [12] that is selectively toxic to cancer cells, including multidrug-resistant cancer cell lines [13–17]. Annonaceous acetogenins induce cytotoxicity by inhibiting the mitochondrial complex I, which is involved in ATP synthesis [14]. As cancer cells have a higher demand for ATP than the normal cells, mitochondrial complex I inhibitors have potential in cancer therapeutics.
A few in vivo studies involving Annona Muricata have been reported. Among these, two reports have shown the ability of the leaf extract to regenerate pancreatic islet β cells in diabetic rats [18, 19]. These studies suggest an additional benefit of the natural product against PC given that diabetes has been classified as a risk factor of the malignant disease [20]. More recently, one study analyzing the anti-tumor efficacy of Annona Muricata was published [21]. The extract had a direct anti-tumorigenic effect on breast cancer cells by downregulating the expression of the epidermal growth factor receptor (EGFR). Although this study demonstrates the potential anti-tumorigenic properties of Graviola, the doses used in the experimental design were not properly controlled. The mice were fed with the extract mixed in the diet and the exact amount ingested by each animal could not be estimated accurately.
Although a few in vitro reports have shown the cytotoxic characteristics of Graviola against various cancer cell lines, including PC cells [12], the comprehensive in vivo effects and mechanistic scientific studies are still lacking. To our knowledge, the studies reported herein are the first to indicate that Graviola extract has promising characteristics for PC therapeutics. Comprehensive in vitro and in vivo studies in various PC cell lines revealed that the natural product inhibited multiple signaling pathways that regulate metabolism, cell cycle, survival, and metastatic properties of PC cells.
Graviola supplement capsules were purchased from Raintree (Carson City, NV). The capsules consisted of 100% pure, finely milled Graviola leaf/stem powder with no binders or fillers. The capsule contents were suspended in DMSO (100mg/mL). After incubating for 5min, the suspension was centrifuged and the supernatant (i.e. extract) was filtered to remove any remaining particles. Subsequent dilutions were prepared in Dulbecco’s modification of Eagle’s medium (DMEM) supplemented with 10% of fetal bovine serum (FBS). Stock solutions and respective dilutions were freshly prepared prior to treatment.
2.2 Cell Culture
The metastatic PC cell lines FG/COLO357 and CD18/HPAF were purchased from the American Type Culture Collection (ATCC). Before performing experiments, the PC cell lines were authenticated by short tandem repeat analysis. It was ensured that PC cells were used at fewer than 20 passages after purchase from ATCC. Cells were cultured in DMEM medium supplemented with 10% FBS and antibiotics (100μg/mL penicillin and 100μg/mL streptomycin). The cells were maintained at 37°C and 5% CO2 in a humidified atmosphere.
2.3 Antibodies
The antibodies for phospho-ERK1/2, total ERK, phospho-Akt (Ser 473), total Akt, NF-κB, and caspase-3 were purchased from Cell Signaling Technology (Danvers, MA). The antibodies for Cyclin-D1, phospho-FAK (Tyr 925), and total FAK were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). The β-actin and β-Tubulin antibodies were obtained from Sigma Aldrich (St. Louis, MO), whereas the HIF-1α antibody was purchased from BD Biosciences (San Jose, CA). The MUC4 monoclonal antibody (8G7) used in these studies was developed by our group [22]. MMP9 antibody was obtained from a hybridoma cell supernatant kindly provided by Dr. Rakesh Singh at UNMC. The secondary antibodies used for western blot analyses were the ECL™ anti-mouse and anti-rabbit IgG conjugated to horseradish peroxidase (GE healthcare, UK). Fluorescein isothiocyanate (FITC) conjugated-anti-mouse and Alexa Fluor conjugated anti-mouse antibodies were obtained from Invitrogen (Carlsbad, CA).
2.4 Cytotoxicity Assay
To determine the cytotoxicity of Graviola extract on PC cells, 1×104 cells were seeded per well on a 96-well plate in DMEM supplemented with 10% FBS and antibiotics. After overnight incubation, different concentrations (10–200μg/mL) of the extract were added into triplicate wells. After 48hr, the media was replaced with fresh media containing thiazolyl blue tetrazolium bromide (MTT) reagent (Sigma Aldrich, St. Louis, MO). After 4hr incubation at 37°C in 5% CO2 in humidified atmosphere, the media was replaced with 100μL of DMSO and the corresponding cytotoxicity values were calculated (λ=540nm). The experiment was repeated at least three times.
2.5 Western Blot Analysis
For protein analysis, 0.5×106 of PC cells were seeded on each well of a six-well plate in DMEM supplemented with 10% FBS and antibiotics. After overnight incubation, fresh solutions of Graviola (0–200μg/mL) were prepared and added to the respective wells. Cells incubated with the corresponding amount of DMSO present in the highest concentrated solution of Graviola were used as a negative control (0μg/mL). After 48hr of incubation with the extract, protein lysates were isolated and prepared for western blot analysis, as previously described [23].
2.6 Real-time PCR
The transcripts levels of the glucose transporters GLUT1 and GLUT4, the glycolytic enzymes hexokinase II (HKII) and lactate dehydrogenase A (LDHA), and the mucin glycoprotein MUC4 in PC cells were determined after treatment with Graviola extract by real-time PCR. 0.5×106 cells were seeded in each well of a six-well plate in complete media. After overnight incubation, fresh solutions of Graviola extract (50 and 100μg/mL) were prepared and cells were incubated for 48hr. Subsequently, cDNA was synthesized from purified RNA and real-time PCR was carried out as has been described by previous studies [23]. The sequences of the gene-specific primers used were: GLUT1: F 5′-GCCATGGAGCCCAGCAGCAA-3′; R 5′-CGGGGACTCTCGGGGCAGAA-3′ GLUT4: F 5′-GCCTGTGGCCACTGCTCCTG-3′; R 5′-GGGGTCTCTGGGCCGGGTAG-3′ HKII: F 5′-GTCATCCCCTTGTGTCAGAG-3′; R 5′-CTTCATTAGTGTCCCCATCCTG-3′ LDHA: F 5′-CCAGTGTGCCTGTATGGAGTG-3′; R 5′-GCACTCTCAACCACCTGCTTG-3′ MUC4: F 5′-GTGACCATGGAGGCCAGTG-3′; R 5′-TCATGCTCAGGTGTCCACAG-3′
2.7 Glucose Uptake
Glucose-uptake rate was assayed by utilizing [3H] 2-deoxyglucose ([3H] 2-DG). 5×104 PC cells were seeded per well in a 24-well plate. 12hr later, the cells were treated with Graviola extract (10 and 50μg/mL) for 48hr. The cells were then starved for glucose for 2hr and incubated for 20min with 2 Ci [3H] 2-DG. Subsequently, cells were lysed with 1% SDS and the lysates were counted for [3H] by utilizing a scintillation counter. Cells treated with labeled and excess unlabeled 2-DG were used as controls to set a baseline for non-specific [3H] uptake. The results were normalized to the cell counts for treated and untreated groups. Glucose uptake was normalized with that of the control cells (0μg/mL) and it is presented as the mean values ± standard error from experiments performed in triplicate.
2.8 ATP Quantification
The CellTiter-Glo® Luminescent Cell Viability Assay (Promega, Madison, WI) was used to measure the ATP content in the cells. Briefly, 1×104 PC cells were seeded in each well of an opaque 96-well plate. Cells were seeded for both ATP quantification and protein concentration estimation. Starting the next day, the cells were incubated with Graviola extract-containing media for 48hr. Subsequently, the instructions of the manufacturer for ATP quantification were followed and luminescence was measured on a Synergy™Mx Luminescent Plate Reader (BioTek, Winooski, VT). Data is presented as the mean value for samples in triplicates, normalized with the protein content for each treatment, as determined by utilizing micro-BCA protein estimation kit.
2.9 Detection and Quantification of Apoptosis and Necrosis
To quantify the number of PC cells undergoing apoptosis and necrosis after being incubated with Graviola extract, the Annexin-V-FLUOS staining kit (Roche Diagnostics, Indianapolis, IN) was used. PC cells were seeded and treated with Graviola extract as described above. After 48hr of treatment with Graviola extract, the instructions of the manufacturer were followed for staining cells for flow cytometry analysis. The experiment was repeated three times.
2.10 Cell Cycle Analysis
PC cells were synchronized at the G1/S phase using a double thymidine block. After seeding cells in 100cm2 Petri dishes, thymidine (2mM) was added for 12hr. After washing cells with serum-free media, the cells were released from thymidine block by culturing in fresh medium containing 24mM 2-deoxycytidine for 9hr. Then, cells were washed and incubated once more with thymidine (2mM) for 14hr. Subsequently, the cells were released from the second thymidine block and the respective treatment prepared in complete media was added for 48hr. For cell cycle analysis, cells were trypsinized and washed with PBS after the duration of the treatment. Cells were then fixed in 70% ethanol at 4°C for 1hr. After washing, cells were incubated with Telford reagent (EDTA, RNAse A, propidium iodide, Triton X-100 in PBS) at 4°C and analyzed by flow cytometry on the next day.
2.11 Confocal Microscopy
For confocal analysis, 2×105 PC cells were seeded on sterilized round glass cover slips. After overnight incubation, Graviola extract (0, 50 and 100μg/mL) was added to the cells, followed by a 48hr incubation. For the detection of reactive oxygen species (ROS), Graviola extract-treated PC cells were incubated with 1μM 2′-7′-Dichlorofluorescein diacetate (DCFH-DA) (Sigma Aldrich, St. Louis, MO) for 15 min. After three washes with PBS, glass cover slips were mounted on glass slides and visualized by confocal microscopy. For β-tubulin and MUC4 confocal analysis, details of the procedure are published elsewhere [23]. Finally, to visualize the arrangement of actin filaments in Graviola extract-treated cells, the cells were stained with fluorescent phallotoxins (Invitrogen, Carlsbad, CA). The instructions of the manufacturer were followed for formaldehyde-fixed cells. Post-staining, the glass cover slips were mounted with Vectashield medium (Vector Laboratories, Burlingame, CA). LSM 510 microscope, a laser scanning confocal microscope (Carl Zeiss GmbH, Thornwood, NY) was utilized to image the cells in the respective channels at a magnification of 630X.
2.12 Wound Healing Assay
For wound healing assays, 3×106 of PC cells were seeded in 60mm petri dishes in DMEM media supplemented with 10% FBS and antibiotics. After overnight incubation, an artificial wound was induced on 100% confluent PC cell monolayers using a sterile pipette tip. Graviola extract-containing (0, 50, 100μg/mL) media solutions were then added to the respective treatment plate. Images (40X) were captured immediately after adding Graviola extract (0hr) and after 24hr of treatment, by a light microscope. The motility of the cells across the wound was visualized in each treatment group.
2.13 Motility Assay
The effect of Graviola extract on the migration of PC cells was also analyzed by a transwell migration assay. FG/COLO357 cells (0.5×106) were suspended in Graviola extract-containing (0–100μg/mL) 1% FBS-DMEM media and seeded for 48hr in 8μm pore size polyethylene terephthalate (PET) membranes (Becton Dickinson, San Jose, CA). DMEM supplemented with 10% FBS was added at the bottom of each well and after 48hr of incubation, the cells that migrated to the bottom of the PET membrane were stained with Diff-Quick cell staining kit (Dade Behring Inc., Newark, DE). The number of cells migrated was quantified by performing cell counts of 10 random fields at 100X magnification. The results are presented as the average number of cells in one field.
2.14 In vivo tumorigenicity studies
The effect of Graviola extract on pancreatic tumor growth was evaluated on orthotopic tumor xenografts. 6–8 week old female athymic immunodeficient mice were purchased from the Animal Production Area of the NCI/Frederick Cancer Research and Development Center (Frederick, MD). The mice were treated in accordance with the Institutional Animal Care and Use Committee (IACUC) guidelines at UNMC and were housed in pathogen-free environment and were fed sterile water and food ad libitum.
Over 90% viable luciferase-labeled CD18/HPAF cells transduced with retroviral particles (Addgene, Cambridge, MA) were orthotopically injected into the head of the pancreas of immunodeficient mice. Details of the orthotopic implantation procedure are described elsewhere [22, 24]. After 1 week of tumor growth, oral gavage treatment of PBS-suspended Graviola extract was given daily for 35 days. The doses of Graviola extract for these studies were based on previous in vivo studies [18, 19, 25] and on the recommended dose for human consumption [11]. Treatment groups (N=8) included: PBS only (0 mg/kg), 50mg/kg, and 100mg/kg Graviola extract. Graviola extract was not dissolved in DMSO for these studies in order to demonstrate the benefit of the aqueous natural oral supplement in PC therapy. Nevertheless, the cytotoxic properties of the Graviola extract suspended in PBS were corroborated beforehand (Supplementary Fig. 1). In vivo IVIS 200 biophotonic imaging system was used to capture images (Caliper Life Sciences, Hopkinton, MA) of pancreatic tumors within every two weeks during the course of treatment with Graviola extract. Mice were sacrificed after 42 days of tumor growth and 35 days of treatment with Graviola extract. Changes in tumor growth and sites of metastasis were evaluated in each treatment group. Body weights of mice were measured before the treatment.
2.15 Analysis of pancreatic tumor tissues
On the necropsy day, pancreatic tumors from the different treatment groups were divided for protein and immunohistochemistry (IHC) analyses. The tumors were immediately frozen under liquid nitrogen for protein analysis. To prepare tumor lysates, the tumors were then suspended on radioimmunoprecipitation (RIPA) buffer and sonicated for three cycles with a Branson digital sonifier® (60% amplitude, 10s). After centrifuging the homogeneous suspension, the protein concentration in each sample was estimated and respective solutions for western blot analyses were prepared as previously described [23].
For histopathological and IHC analyses, the tumor tissues were fixed in 10% Formalin for 48hr. The tumors were embedded in paraffin and 5μm sections were cut and stained with hematoxylin and eosin stains (H&E) and various antibodies (i.e. MMP9 and MUC4). Details of the procedure for IHC staining is described elsewhere [24]. The IHC and H&E stained slides were evaluated by pathologist at University of Nebraska Medical Centre.
2.16 Statistical Analysis
The JMP® Statistical Discovery Software (Cary, NC) was used to determine the statistical significance within the treatment replicates in each experiment. A Student’s t-test was used to calculate the corresponding p-value. All p values < 0.05 were considered statistically significant.
3.1 Graviola extract induces cytotoxicity of pancreatic cancer cells
The PC cells FG/COLO357 and CD18/HPAF were incubated for 48hr with different concentrations of Graviola extract. The results from the MTT cytotoxicity assay indicated a progressive decrease in cell viability with the successive increase in the concentrations of the extract (Fig. 1A). After 48hr of treatment, the resulting IC50 of Graviola extract on FG/COLO357 and CD18/HPAF cells was 200 and 73μg/mL, respectively (Fig. 1B) and the results indicated that CD18/HPAF cell line is more sensitive to the Graviola extract than the FG/COLO357 cell line.
Effect of Graviola extract on pancreatic cancer cell viability
It is well known that the activation of the extracellular signal-regulated kinase (ERK) and the phosphatidylinositol 3′kinase (PI3K/Akt) pathways play a crucial role in the proliferation and survival of PC [26] and inhibition of these pathways leads to the inhibition of pancreatic tumor growth [27, 28]. The present study revealed that treatment of PC cells with Graviola extract resulted in decreased activation of both ERK and Akt pathways in PC cells (Fig. 1C). Thus, the inhibition of these pathways is in agreement with the decreased viability of PC cells treated with Graviola extract.
3.2 Pancreatic cancer cell metabolism is inhibited by Graviola extract
Previous studies have shown that major bioactive components present in Graviola extract that inhibit mitochondrial complex I [13–17], suggesting their direct involvement in cell metabolism. It has already been well-documented that cancer cells undergo a metabolic shift to adapt and survive under harsh environments by enhancing aerobic glycolysis [29, 30]. Also, Akt activation leads to glycolytic ATP generation in tumor cells [31]. Hence, the effect of Graviola extract on several stages of the glycolytic pathway in PC cells was analyzed.
The expression of HIF-1α, a critical regulator of aerobic glycolysis in cancer cells [32], was analyzed in PC cells after incubation with Graviola extract (Fig. 2A). We observed reduced HIF-1α expression in both PC cell lines, suggesting a direct effect of this natural product on the metabolism of PC cells. Likewise, it has been previously reported that the NF-κB upregulates the expression of HIF-1α [33, 34]. Not surprisingly, the expression levels of NF-κB were also reduced in PC cells after being incubated with Graviola extract (Fig. 2A).
Effect of Graviola extract on the metabolism of pancreatic cancer cells
Subsequently, the expression of the glucose transporters 1 and 4 (GLUT1 and GLUT4), and the expression of the glycolytic enzymes hexokinase II (HKII) and lactate dehydrogenase A (LDHA), all of which are upregulated by HIF1-α in cancer cells [32, 35], were analyzed in Graviola extract-treated PC cells by real-time PCR analysis (Fig 2B). Overall, the transcript levels of GLUT1, HKII, and LDHA were significantly reduced in both PC cell lines when compared to untreated cells (i.e. 60–87% downregulation).
Cancer cells have an increased expression of glucose transporters to enhance glucose uptake, which in turn increases the glycolytic rate for an enhanced ATP production that will ultimately lead to an enhanced tumor growth [10]. Thus, based on the results discussed above, it was not surprising that PC cells treated with Graviola extract doses over 10 μg/mL had a decreased rate of glucose uptake when compared to untreated cells (0μg/mL) (Fig. 2C). Finally, to evaluate the energy outcome of the glycolytic pathway of PC cells, we measured ATP production in Graviola extract-treated PC cells (Fig. 2D), and observed significant inhibition by 42–47% and by 31–43% doses in FG/COLO357 and CD18/HPAF PC cells, respectively. Altogether, these results indicate that Graviola extract impairs the metabolism of PC cells that will ultimately lead to decreased cell viability.
3.3 Graviola extract induces necrosis of pancreatic cancer cells
In order to evaluate the cytotoxic pathways induced by Graviola extract. PC cells were stained with annexin-V and propidium iodide (PI) staining to measure the necrotic and apoptotic cell populations by performing flow cytometry. While the necrotic cell population in both PC cell lines increased significantly after incubation with Graviola extract, the apoptotic cell population remained unchanged (Fig. 3A). Subsequently, the production of Graviola extract-induced reactive oxygen species (ROS) in FG/COLO357 and CD18/HPAF PC cells was confirmed by confocal microscopy (Fig. 3B). Additionally, it was also observed that cells incubated with Graviola extract have a gain in cell volume, a characteristic of necrotic cell death.
Analysis of cytotoxic mechanism of Graviola extract in pancreatic cancer cells
In order to confirm that Graviola extract was not inducing apoptosis of PC cells, the levels of caspase-3 expression were analyzed by western blot analysis. The Caspase-3 expression values remained statisticallyunaltered by treatment with the extract, suggesting that apoptotic pathways are not involved (Fig. 3C). Furthermore, the apoptotic cells population in Graviola extract-treated cells was also analyzed by Telford staining, and the results corroborated the findings from AnnexinV/PI staining studies, where the number of apoptotic cell population did not vary after being incubated with the natural compound (data not shown).
An analysis of the different phases of the cell cycle after treatment with Graviola extract demonstrated cell cycle arrest at G1 phase (Fig. 3D). While the G1 cell population increased from 43 to 65%, the S phase decreased from 56 to 32% with increasing concentrations of Graviola extract (0, 5, 100μg/mL). To support these results, the expression of CyclinD1 in Graviola extract-treated PC cells was analyzed (Fig. 3E). In agreement with previous studies indicating that a decreased CyclinD1 expression induces G0/G1 cell cycle arrest [36], Graviola extract-treated PC cells had also reduced expression of the cell cycle regulatory protein.
3.4 Motility of pancreatic cancer cells decreases after treatment with Graviola extract
The effect of Graviola extract on the functional properties of PC cells was analyzed in vitro wound healing and migration assays (Fig. 4A, B). As it can be observed in the images from the wound healing assays, PC cells treated with Graviola extract did not close the wound even after 24hr, as opposed to untreated cells (0μg/mL), indicating reduced motility of PC cells after treatment with Graviola extract (Fig. 4A). Similarly, the migratory capacity of PC cells was also reduced after treatment with Graviola extract, as evaluated by a transwell assay (Fig. 4B), suggesting that the natural extract reduces the motility of PC cells.
Effect of Graviola extract in the motility, migration, and cytoskeleton of pancreatic cancer cells
The motility and migration of cancer cells is associated with the rearrangements of the cortical actin and microtubules network [37, 38]. Additionally, cellular ATP depletion has been associated with reorganization of the actin cytoskeleton [39] and suppression of the dynamics of microtubules is known to induce mitotic arrest [40]. Taking this into consideration, the cytoskeleton of Graviola extract-treated PC cells was analyzed by confocal microscopy (Fig. 4C, D). The image results of phallotoxins (i.e. phalloidin) staining indicate a disruption of the cortical actin network and dissolution of stress fibers in Graviola extract-treated PC cells (Fig. 4C). Similarly, a disruption of microtubules dynamics was evident after β-tubulin staining of PC cells incubated with Graviola extract (Fig. 4D). To further analyze the effect of Graviola extract on motility and migration of PC cells, the expression levels of the phosphorylated focal adhesion kinase (pFAK), which is involved in mitogenic signaling and motility [41], and matrix metalloproteinase 9 (MMP9), which targets many extracellular proteins including adhesion molecules [42], were analyzed by western blot analysis (Fig. 4E). In agreement with the experiments discussed above, we observed that the expression levels of both pFAK and MMP9 were downregulated in Graviola extract-treated cells.
3.5 Graviola extract inhibits tumor growth and metastasis of pancreatic cancer cells
Based on the results obtained from in vitro experiments, Graviola extract has promising properties to be incorporated in PC therapeutics. Nevertheless, these anti-tumorigenic properties require further validation through in vivo experiments. In order to evaluate the therapeutic potential of Graviola extract, a more realistic situation for administering the extract was mimicked. It is recommended that Graviola extract supplement must be taken on a regular basis [11], and therefore, it was decided that the extract must be administered by oral gavage after suspending contents of the capsule in aqueous solution instead of dissolving it in DMSO. Prior to evaluating the anti-tumorigenic properties of aqueous Graviola extract suspension by in vivo experiments, pertinent in vitro experiments corroborating the cytotoxic potential of the aqueous suspensions on PC cells were completed beforehand (Supplementary Fig. 1).
For tumorigenic studies, CD18/HPAF cells expressing luciferase were orthotopically injected into the pancreas of athymic mice. After 1 week, in vivo biophotonic imaging confirmed tumor growth in all animals and the treatment regimen was initiated. The tumor growth during the treatment was monitored by imaging every two weeks. After 35 days of treatment, the animals were euthanized and the pancreatic tumors were removed and weighed. Although pancreatic tumors were not completely eradicated, the results indicate that tumor growth decreased significantly in Graviola extract-treated mice in comparison to the control group (Fig. 5A). Specifically, the tumor growth inhibition in mice treated with a dose of 50mg/kg Graviola extract was 59.8% (p-value=0.0008) whereas in mice treated with 100 mg/kg Graviola extract the inhibition was 50.3% (p-value = 0.006), indicating the efficacy of the natural product in PC regression. The metastatic lesions in each mouse were evaluated in various vital organs including the liver, spleen, mesenteric lymph nodes (LN), small and large intestines, peritoneum, diaphragm, and ovaries (Fig. 5B). Although all the metastatic lesions were reduced in Graviola extract-treated mice in comparison to the untreated control mice, the incidence of metastasis in the liver, mesenteric LN, and ovaries was significantly reduced (p-values ≤ 0.02). Representative biophotonic tumor images illustrate the tumor growth across the different groups during the course of the treatment (Fig. 5C).
Evaluation of Graviola Extract in pancreatic cancer orthotopic xenograft model
Further, tumors were evaluated by H&E (Fig. 5D) and IHC staining (Fig. 6). The H&E stained tumor sections showed necrotic cells in 20–50% of the pancreatic tumor tissues from Graviola extract-treated mice as compared with tumors from the control mice. These results further strengthen the results from in vitro experiments, which demonstrate that Graviola extract-mediated reduction in PC cell viability was through the induction of necrosis.
Immunohistochemical analyses of pancreatic tumors after treatment with Graviola extract
The tumor lysates and paraffin embedded pancreatic tumors were also evaluated by IHC for the expression of MMP9 (Fig. 6A) and MUC4 (Fig. 6B). In agreement with in vitro data, the levels of MMP9 were reduced in tumors from Graviola extract-treated mice compared to the untreated controls. As the expression of MMP9 has been related to invasion and metastasis, the reduced levels of the protein in Graviola extract-treated tumors substantiate our findings of reduced metastatic sites in these mice.
Previous studies performed by our group have established the correlation of the expression of mucin4 (MUC4) glycoprotein with progression and metastasis of PC [24, 43–45]. Therefore, we were particularly interested in evaluating the effect of Graviola extract on the expression of MUC4 in PC cells and pancreatic tumors. In vitro experiments demonstrated a significant downregulation in MUC4 expression, both at the translational (Supplementary Figs. 1C, 2A, B) and transcriptional levels (Supplementary Fig. 2C) in Graviola extract treated PC cells. Similarly, the expression of the MUC4 was reduced in pancreatic tumors from mice treated with Graviola extract as compared to the untreated mice (Fig. 6B), Further supporting our findings of reduced tumor growth and metastasis after treatment with Graviola extract.
Little or no progress has been accomplished in PC treatment over the last 40 years. Novel therapeutics against this lethal malignancy must inhibit several pathways that promote survival, progression, and metastasis of PC cells. Based on the fact that cancer cells are mainly dependent on the glycolytic pathway for ATP production, glucose deprivation by anti-glycolytic drugs can induce cancer cell death [46], a pathway that can be targeted and explored in PC therapies [47].
Natural products have been investigated in PC therapeutics over several decades, but to date none has been incorporated in routine chemotherapies [10]. Traditionally, the leaves from Graviola (Annona Muricata) have been used for a wide range of human diseases including cancer [11]. The present study is the first to demonstrate that Graviola extract reduces the viability of PC cells and tumors by inducing necrosis and cell cycle arrest, and by inhibiting PC cell motility (i.e. cytoskeleton rearrangement), migration, and metabolism. Overall, in vitro experiments revealed that the compounds present in the natural extract inhibited several pathways involved in PC cell proliferation and metabolism, simultaneously. Such inhibitions ultimately led to a decrease in tumor growth and metastasis in orthotopically transplanted pancreatic tumor-bearing mice.
In PC patients, an increased metabolic activity and glucose concentration of malignant tumors has been linked to pancreatic tumor aggressiveness [47]. Additionally, the presence of hypoxia in PC has been associated with tumor growth and metastasis [48, 49]. Indeed, the presence of hypoxic environment has been linked to the oncogenic and metabolic transformation (i.e. glycolysis) of PC cells that results in resistance to conventional cancer therapeutics [48, 50]. More specifically, it has been suggested that hypoxia can induce resistance to gemcitabine through the activation of PI3K/Akt/NF-κB and MAPK/ERK pathways [51], which are also related to PC progression and survival. The activation of both of these signaling pathways was evaluated in PC cells after treatment with Graviola extract and it was found that the extract suppressed phosphorylation of the key molecules involved in these pathways, which correlated with reduced viability of PC cells. Subsequently, the expression of HIF-1α, the major transcription factor activated under hypoxic conditions, and its ensuing downstream effects on PC cell metabolism were analyzed in Graviola extract-treated cells. The results indicated the natural product inhibited PC cell metabolism by inhibiting the expression of HIF-1α, NF-κB, glucose transporters (i.e. GLUT1, GLUT4), and glycolytic enzymes (i.e. HKII, LDHA), all of which lead to the reduction of glucose uptake and ATP production by PC cells.
The overall downregulation of PC cell metabolism induced by Graviola extract resulted in PC cell death and necrosis. In agreement with previous studies of ATP reduction, the metabolic and therapeutic stress induced by Graviola extract led to an acute ATP depletion, which is accompanied by increased intracellular ROS, ultimately leading to necrosis [52–54]. While necrotic agents have not been considered beneficial in cancer therapies due to induction of local inflammation, the process itself can lead to the activation of the innate immune system capable of initiating anti-tumor immunity [52]. It makes it imperative to evaluate the effect of a necrosis-inducing product such as Graviola extract in an immune competent host. In this regard, we plan to evaluate the effect of the natural product on the progression of pancreatic adenocarcinoma in the KrasG12DPdx1-Cre spontaneous animal model, where the effect on the immune system can be evaluated.[55, 56]. In order to evaluate the potential of Graviola extract in preventing PC progression, we plan to supplement the diet of KrasG12DPdx1-Cre mice with Graviola extract after the mice start developing pancreatic intraepithelial neoplastic (PanIN) lesions. The effective concentrations of Graviola metabolites after oral absorption and effects on the immune system will be measured as well. Additional experiments will be carried out to evaluate the potential of a combination therapy of Graviola extract with the standard chemotherapeutic drug Gemcitabine. With the results discussed in the present study, it is expected that minimum doses of the chemotherapeutic drug will be needed to eradicate the malignant disease.
The major bioactive compounds identified in Annona Muricata have been classified as Annonaceous acetogenins, which inhibit mitochondrial complex I that leads to a decreased ATP production [13–17]. Although the natural extract capsules used in these studies contained numerous compounds, the presence of Annonaceous acetogenins was evident by the depletion of ATP production in PC cells after being incubated with Graviola extract. Bioactivity-guided fractionation for the identification of potent bioactive (i.e. anti-tumorigenic) compounds that are present in the Graviola extract is currently being investigated. We are also ensuring that cytotoxic effects are specific to tumorigenic cells only, by including the non-transformed immortalized pancreatic epithelial cell line HPNE, which is derived from pancreatic duct (data not shown).
Pancreatic tumors develop from a complex interplay of numerous signaling pathways and Graviola extract has shown promising anti-tumorigenic characteristics by targeting some of these pathways all at once. Although novel glycolytic inhibitors, such as Graviola extract, may have broad therapeutics applications [57], inhibition of glycolysis alone may not be sufficient to eradicate tumor cells completely. Perhaps the use of alternative medicine, like taking Graviola capsules on a regular basis, should still be considered a supplement, not a replacement for standard therapies. Currently, in vitro studies evaluating the potential of the natural product in combination with chemotherapeutic drugs are being conducted.
Supplementary Figure 1 In vitro evaluation of aqueous solutions of Graviola extract in pancreatic cancer cells:
(A) Cytotoxicity of Graviola extract suspended in PBS. The PC cells CD18/HPAF were incubated with different doses of Graviola extract suspended in PBS for 48hr. Cytotoxicity was analyzed by MTT cytotoxic assay. Data from experiments performed in triplicate is presented as the mean value ± standard error of mean. (*p-value<0.0001, **p-value=0.0017, compared to untreated cells); (B) ATP quantification of CD18/HPAF PC cells after treatment with Graviola extract suspended in PBS. Data from experiments performed in triplicate is presented as the mean value normalized with total protein content ± standard error of mean; (C) Western blot analysis of the expression of proteins related to the proliferation, invasion, and metastasis of PC in CD18/HPAF cells after incubation with Graviola extract suspended in PBS. Protein lysates (30μg) were resolved on 2% SDS agarose gels (MUC4) and 10% SDS-PAGE (HIF-1α, Caspase3, MMP9, and β-actin). β-actin was used as a loading control.
Supplementary Figure 2 Expression of MUC4 glycoprotein in pancreatic cancer cells after treatment with Graviola extract:
(A) Western blot analysis of MUC4 and HER2 in PC cells after treatment with Graviola extract. Protein lysates (30μg) were resolved on 2% agarose gels for MUC4 and 10% SDS-PAGE for HER2. β-actin was used as a loading control; (B) Confocal microscope images of PC cells stained against MUC4 (anti-Alexa Fluor 594) after treatment with Graviola extract. Cell nuclei were stained with DAPI. Scale bars represent 10μm; (C) Measurement of MUC4 transcripts in PC cells incubated with Graviola extract by real-time PCR. Data is presented as the average fold difference of MUC4 levels in Graviola extract-treated cells versus untreated cells (0μg/mL) ± standard error of mean. The housekeeping gene β-actin was used as an internal control. (*p-value<0.005; **p-value<0.005, compared to untreated PC cells)
The invaluable technical support from Kavita Mallya is greatly appreciated. We would like to give special thanks to UNMC professors: Dr. Michel Ouellette for kindly providing CD18/HPAF-Luciferase and HPNE cells, Dr. Shilpa Buch for allowing us to use the Luminescence plate reader, Dr. Vimla Band for allowing us to use the microscope to image tumor H&E and IHC sections, and Dr. Steve Caplan for assisting with the analysis of confocal images and providing us the β-Tubulin antibody. We also thank Janice A. Tayor and James R. Talaska of the Confocal Laser Scanning Microscope Core Facility at UNMC, Victoria B. Smith and Megan Michalak of the UNMC Cell Analysis Core Facility, and Ms. Kristi Berger, the Eppley Cancer Center for editing this manuscript. We are also very grateful for the expertise and involvement of Drs. Amarnath Natarajan and Abijah Nyong from the Chemistry department at UNMC in the bioactivity-guided fractionation of the Graviola extract. The authors of this work are supported by grants from the National Institutes of Health: NIH-NCI Cancer Biology Training Grant UNMC T32CA009479, R01 CA78590, U01EDRN CA111294, R01 CA131944, R01 CA133774, R01 CA 138791, P50 SPORE CA127297 and U54 CA160163.
There are no potential conflicts of interest involved with this work.
Publisher’s Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
1. SEER Stat Fact Sheets:Pancreas. National Cancer Institute; Oct 28, 2011.
2. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA Cancer J Clin. 2012;62:10–29. [PubMed]
3. Chakraborty S, Baine MJ, Sasson AR, Batra SK. Current status of molecular markers for early detection of sporadic pancreatic cancer.Biochim Biophys Acta. 2011;1815:44–64. [PMC free article] [PubMed]
4. Bilimoria KY, Bentrem DJ, Ko CY, Stewart AK, Winchester DP, Talamonti MS. National failure to operate on early stage pancreatic cancer.Ann Surg. 2007;246:173–180. [PMC free article] [PubMed]
5. Bilimoria KY, Bentrem DJ, Ko CY, Tomlinson JS, Stewart AK, Winchester DP, Talamonti MS. Multimodality therapy for pancreatic cancer in the U.S.: utilization, outcomes, and the effect of hospital volume. Cancer. 2007;110:1227–1234. [PubMed]
6. Smith BD, Smith GL, Hurria A, Hortobagyi GN, Buchholz TA. Future of cancer incidence in the United States: burdens upon an aging, changing nation. J Clin Oncol. 2009;27:2758–2765. [PubMed]
7. Burris HA, III, Moore MJ, Andersen J, Green MR, Rothenberg ML, Modiano MR, Cripps MC, Portenoy RK, Storniolo AM, Tarassoff P, Nelson R, Dorr FA, Stephens CD, Von Hoff DD. Improvements in survival and clinical benefit with gemcitabine as first-line therapy for patients with advanced pancreas cancer: a randomized trial. J Clin Oncol. 1997;15:2403–2413. [PubMed]
8. Conroy T, Desseigne F, Ychou M, Bouche O, Guimbaud R, Becouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de la Fouchardiere C, Bennouna J, Bachet JB, Khemissa-Akouz F, Pere-Verge D, Delbaldo C, Assenat E, Chauffert B, Michel P, Montoto-Grillot C, Ducreux M. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N Engl J Med. 2011;364:1817–1825. [PubMed]
9. Jansen RJ, Robinson DP, Stolzenberg-Solomon RZ, Bamlet WR, de Andrade M, Oberg AL, Hammer TJ, Rabe KG, Anderson KE, Olson JE, Sinha R, Petersen GM. Fruit and vegetable consumption is inversely associated with having pancreatic cancer. Cancer Causes Control.2011;22:1613–1625. [PMC free article] [PubMed]
10. Stan SD, Singh SV, Brand RE. Chemoprevention strategies for pancreatic cancer. Nat Rev Gastroenterol Hepatol. 2010;7:347–356.[PMC free article] [PubMed]
11. Taylor L. Herbal Secrets of the Rainforest. 2 Sage Press, Inc; 2002. Technical Data Report for Graviola: Annona Muricata.
12. Kim GS, Zeng L, Alali F, Rogers LL, Wu FE, Sastrodihardjo S, McLaughlin JL. Muricoreacin and murihexocin C, mono-tetrahydrofuran acetogenins, from the leaves of Annona muricata. Phytochemistry. 1998;49:565–571. [PubMed]
13. Oberlies NH, Jones JL, Corbett TH, Fotopoulos SS, McLaughlin JL. Tumor cell growth inhibition by several Annonaceous acetogenins in an in vitro disk diffusion assay. Cancer Lett. 1995;96:55–62. [PubMed]
14. McLaughlin JL. Paw paw and cancer: annonaceous acetogenins from discovery to commercial products. J Nat Prod. 2008;71:1311–1321.[PubMed]
15. Tormo JR, Royo I, Gallardo T, Zafra-Polo MC, Hernandez P, Cortes D, Pelaez F. In vitro antitumor structure-activity relationships of threo/trans/threo mono-tetrahydrofuranic acetogenins: correlations with their inhibition of mitochondrial complex I. Oncol Res. 2003;14:147–154. [PubMed]
17. Liaw CC, Chang FR, Lin CY, Chou CJ, Chiu HF, Wu MJ, Wu YC. New cytotoxic monotetrahydrofuran annonaceous acetogenins from Annona muricata. J Nat Prod. 2002;65:470–475. [PubMed]
18. Adewole SO, Caxton-Martins EA. Morphological Changes and Hypoglycemic Effects of Annona Muricata Linn. (Annonaceae) Leaf Aqueous Extract on Pancreatic B- Cells of Streptozotocin-Treated Diabetic Rats. African J Biomed Res. 2006;9:173–180.
19. Adeyemi DO, Komolafe OA, Adewole SO, Obuotor EM, Abiodum AA, Adenowo TK. Histomorphological and morphometric studies of the pancreatic islet cells of diabetic rats treated with extracts of Annona Muricata. Folia Morphol. 2010;69:92–100. [PubMed]
20. Magruder JT, Elahi D, Andersen DK. Diabetes and pancreatic cancer: chicken or egg? Pancreas. 2011;40:339–351. [PubMed]
21. Dai Y, Hogan S, Schmelz EM, Ju YH, Canning C, Zhou K. Selective growth inhibition of human breast cancer cells by graviola fruit extract in vitro and in vivo involving downregulation of EGFR expression. Nutr Cancer. 2011;63:795–801. [PubMed]
22. Moniaux N, Varshney GC, Chauhan SC, Copin MC, Jain M, Wittel UA, Andrianifahanana M, Aubert JP, Batra SK. Generation and characterization of anti-MUC4 monoclonal antibodies reactive with normal and cancer cells in humans. J Histochem Cytochem. 2004;52:253–261.[PubMed]
23. Torres MP, Ponnusamy MP, Chakraborty S, Smith LM, Das S, Arafat HA, Batra SK. Effects of thymoquinone in the expression of mucin 4 in pancreatic cancer cells: implications for the development of novel cancer therapies. Mol Cancer Ther. 2010;9:1419–1431. [PMC free article][PubMed]
24. Singh AP, Moniaux N, Chauhan SC, Meza JL, Batra SK. Inhibition of MUC4 expression suppresses pancreatic tumor cell growth and metastasis. Cancer Res. 2004;64:622–630. [PubMed]
25. de Sousa OV, Vieira GD, de Jesus RG, Yamamoto CH, Alves MS. Antinociceptive and Anti-Inflammatory Activities of the Ethanol Extract of Annona muricata L. Leaves in Animal Models. Int J Mol Sci. 2010;11:2067–2078. [PMC free article] [PubMed]
26. Seufferlein T. Novel protein kinases in pancreatic cell growth and cancer. Int J Gastrointest Cancer. 2002;31:15–21. [PubMed]
27. Chang Q, Chapman MS, Miner JN, Hedley DW. Antitumour activity of a potent MEK inhibitor RDEA119/BAY 869766 combined with rapamycin in human orthotopic primary pancreatic cancer xenografts. BMC Cancer. 2010;10:515. [PMC free article] [PubMed]
28. Wei WT, Chen H, Ni ZL, Liu HB, Tong HF, Fan L, Liu A, Qiu MX, Liu DL, Guo HC, Wang ZH, Lin SZ. Antitumor and apoptosis-promoting properties of emodin, an anthraquinone derivative from Rheum officinale Baill, against pancreatic cancer in mice via inhibition of Akt activation.Int J Oncol. 2011;39:1381–1390. [PubMed]
29. DeBerardinis RJ, Lum JJ, Hatzivassiliou G, Thompson CB. The biology of cancer: metabolic reprogramming fuels cell growth and proliferation. Cell Metab. 2008;7:11–20. [PubMed]
30. Kroemer G, Pouyssegur J. Tumor cell metabolism: cancer’s Achilles’ heel. Cancer Cell. 2008;13:472–482. [PubMed]
31. Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nat Rev Cancer. 2011;11:85–95. [PubMed]
32. Lu H, Forbes RA, Verma A. Hypoxia-inducible factor 1 activation by aerobic glycolysis implicates the Warburg effect in carcinogenesis. J Biol Chem. 2002;277:23111–23115. [PubMed]
33. Fitzpatrick SF, Tambuwala MM, Bruning U, Schaible B, Scholz CC, Byrne A, O’Connor A, Gallagher WM, Lenihan CR, Garvey JF, Howell K, Fallon PG, Cummins EP, Taylor CT. An intact canonical NF-kappaB pathway is required for inflammatory gene expression in response to hypoxia.J Immunol. 2011;186:1091–1096. [PubMed]
34. Nam SY, Ko YS, Jung J, Yoon J, Kim YH, Choi YJ, Park JW, Chang MS, Kim WH, Lee BL. A hypoxia-dependent upregulation of hypoxia-inducible factor-1 by nuclear factor-kappaB promotes gastric tumour growth and angiogenesis. Br J Cancer. 2011;104:166–174. [PMC free article][PubMed]
35. Ke Q, Costa M. Hypoxia-inducible factor-1 (HIF-1) Mol Pharmacol. 2006;70:1469–1480. [PubMed]
36. Masamha CP, Benbrook DM. Cyclin D1 degradation is sufficient to induce G1 cell cycle arrest despite constitutive expression of cyclin E2 in ovarian cancer cells. Cancer Res. 2009;69:6565–6572. [PubMed]
37. Cunningham CC. Actin structural proteins in cell motility. Cancer Metastasis Rev. 1992;11:69–77. [PubMed]
38. Kaverina I, Straube A. Regulation of cell migration by dynamic microtubules. Semin Cell Dev Biol. 2011 [PMC free article] [PubMed]
39. Bacallao R, Garfinkel A, Monke S, Zampighi G, Mandel LJ. ATP depletion: a novel method to study junctional properties in epithelial tissues. I. Rearrangement of the actin cytoskeleton. J Cell Sci. 1994;107(Pt 12):3301–3313. [PubMed]
40. Jordan MA, Horwitz SB, Lobert S, Correia JJ. Exploring the mechanisms of action of the novel microtubule inhibitor vinflunine. Semin Oncol.2008;35:S6–S12. [PubMed]
41. Zhao X, Guan JL. Focal adhesion kinase and its signaling pathways in cell migration and angiogenesis. Adv Drug Deliv Rev. 2011;63:610–615.[PMC free article] [PubMed]
42. McCawley LJ, Matrisian LM. Matrix metalloproteinases: they’re not just for matrix anymore! Curr Opin Cell Biol. 2001;13:534–540.[PubMed]
43. Bafna S, Kaur S, Momi N, Batra SK. Pancreatic cancer cells resistance to gemcitabine: the role of MUC4 mucin. Br J Cancer. 2009;101:1155–1161. [PMC free article] [PubMed]
44. Chaturvedi P, Singh AP, Moniaux N, Senapati S, Chakraborty S, Meza JL, Batra SK. MUC4 mucin potentiates pancreatic tumor cell proliferation, survival, and invasive properties and interferes with its interaction to extracellular matrix proteins. Mol Cancer Res. 2007;5:309–320. [PubMed]
45. Swartz MJ, Batra SK, Varshney GC, Hollingsworth MA, Yeo CJ, Cameron JL, Wilentz RE, Hruban RH, Argani P. MUC4 expression increases progressively in pancreatic intraepithelial neoplasia. Am J Clin Pathol. 2002;117:791–796. [PubMed]
46. El MN, Caro-Maldonado A, Ramirez-Peinado S, Munoz-Pinedo C. Sugar-free approaches to cancer cell killing. Oncogene. 2011;30:253–264.[PubMed]
47. Komar G, Kauhanen S, Liukko K, Seppanen M, Kajander S, Ovaska J, Nuutila P, Minn H. Decreased blood flow with increased metabolic activity: a novel sign of pancreatic tumor aggressiveness. Clin Cancer Res. 2009;15:5511–5517. [PubMed]
48. Vasseur S, Tomasini R, Tournaire R, Iovanna JL. Hypoxia Induced Tumor Metabolic Switch Contributes to Pancreatic Cancer Aggressiveness.Cancers. 2010;2:2138–2152.
49. Duffy JP, Eibl G, Reber HA, Hines OJ. Influence of hypoxia and neoangiogenesis on the growth of pancreatic cancer. Mol Cancer. 2003;2:12.[PMC free article] [PubMed]
50. Yokoi K, Fidler IJ. Hypoxia increases resistance of human pancreatic cancer cells to apoptosis induced by gemcitabine. Clin Cancer Res.2004;10:2299–2306. [PubMed]
51. Chen EY, Mazure NM, Cooper JA, Giaccia AJ. Hypoxia activates a platelet-derived growth factor receptor/phosphatidylinositol 3-kinase/Akt pathway that results in glycogen synthase kinase-3 inactivation. Cancer Res. 2001;61:2429–2433. [PubMed]
52. Amaravadi RK, Thompson CB. The roles of therapy-induced autophagy and necrosis in cancer treatment. Clin Cancer Res. 2007;13:7271–7279. [PubMed]
53. Eguchi Y, Shimizu S, Tsujimoto Y. Intracellular ATP levels determine cell death fate by apoptosis or necrosis. Cancer Res. 1997;57:1835–1840.[PubMed]
54. Leist M, Single B, Castoldi AF, Kuhnle S, Nicotera P. Intracellular adenosine triphosphate (ATP) concentration: a switch in the decision between apoptosis and necrosis. J Exp Med. 1997;185:1481–1486. [PMC free article] [PubMed]
55. Festjens N, Vanden Berghe T, Vandenabeele P. Necrosis, a well-orchestrated form of cell demise: signalling cascades, important mediators and concomitant immune response. Biochim Biophys Acta. 2006;1757:1371–1387. [PubMed]
56. Golstein P, Kroemer G. Cell death by necrosis: towards a molecular definition. Trends Biochem Sci. 2007;32:37–43. [PubMed]
57. Pelicano H, Martin DS, Xu RH, Huang P. Glycolysis inhibition for anticancer treatment. Oncogene. 2006;25:4633–4646. [PubMed]
Een voordeel van eten volgens Moerman is dat het citroensap en andere vitamine C-rijke producten omvat, evenals andere antioxidantenrijke voeding. Deze antioxidanten maken het nitraat minder schadelijk doordat ze de vorming van nitriet en nitrosamines in het lichaam gedeeltelijk kunnen remmen. Mogelijk kunnen deze stoffen dus de eventuele schadelijke gevolgen van nitriet in het lichaam beperken. Het advies is dus zo min mogelijk bewerkte producten gebruiken, dus geen GECONCENTREERDE bietensap.
Voor de volledigheid: ook andere groenten kunnen rijk zijn aan nitraat, zoals andijvie, bleekselderij, Chinese kool, koolrabi, paksoi, postelein, raapstelen, radijs, rode bieten, sla (alle soorten), spitskool.
Nitraatarme groente:
Asperges, aubergine, bloemkool, boerenkool, broccoli, courgette, doperwten, knolselderij, komkommer, paprika, prei, rodekool, savooiekool, snijbonen, sperziebonen, spruiten, snijbonen, tuinbonen, uien, witlof, wittekool, wortelen.
Nitraat is van nature aanwezig in alle groenten. Nitriet is een stof die ontstaat op basis van nitraat.
Nitraat is niet direct een gevaar voor de gezondheid. Het risico ligt meer in het feit dat het lichaam nitraat om kan zetten in nitriet. Door het bewaren, bereiden of het eten van groente kan nitraat gedeeltelijk omgezet worden in nitriet. Nitriet is sneller schadelijk en nitriet kan weer omgezet worden in het kankerverwekkende nitrosaminen.
Dit zijn schadelijke stoffen die met name ontstaan door het eten van nitraatrijke groenten in combinatie met vis, schaal- en schelpdieren (met uitzondering van zalm en makreel). Door de combinatie van nitriet met de eiwitten afkomstig van de vis ontstaan de schadelijke nitrosaminen.
Uit dierproeven is gebleken dat nitrosaminen mogelijk kankerverwekkend zijn. Zij kunnen zich namelijk binden aan ons erfelijk materiaal (ons DNA) en na verloop van tijd kan dat aanleiding zijn tot tumorvorming.
hansstoop
3 November 2013
Rode Bieten echt superfood.
Ik wil het volgende nogmaals duidelijk onder uw aandacht brengen. Bietensap is een supergezond product dat Cornelis Moerman al zijn patiënten voorschreef. Tegenwoordig noemen we deze producten superfoods.
De rode biet bevat volop voedingsstoffen (fytonutrienten) ze zijn rijk aan vitamine A en C, vezels, ijzer, magnesium, kalium, foliumzuur, betacyanine. Rode bieten zorgen voor een actief afweersysteem en een beschermer tegen kanker. Het is door de eeuwen heen gebruikt tegen o.a. bloedarmoede en nierstoornissen. Het heeft een zuiverende werking op het bloed. Het betacyanide dat voor de diep rode kleur zorgt, samen met andere krachtige antioxidanten, voor het stimuleren van de lever, het remmen van ontstekingen en bieden bescherming tegen huid-, long-, borst-, prostaat-, maag- en darmkanker.De rode biet verlaagt ook het cholesterolgehalte. Wetenschappelijk onderzoek tonen aan dat een hoge bloeddruk te verlagen is door bietensap te drinken en bietensap is tevens een weldaad voor ons hart en onze bloedvaten, maar dat geldt ook voor onze ogen dankzij luteine en zeaxanthine. Naast het vechten tegen kanker, het stimuleren van ons immuunsysteem en het verlagen van onze bloeddruk, gaat het ook dementie tegen.
Jammer dat nu alle aandacht gevestigd wordt op het nitraat, wat van nature in alle groenten zit. Nitraat is n.l. nodig bij het maken van (plantaardige)eiwitten. Echter, we treffen ze meer aan in kasgroenten waarbij kunstmest gebruikt is. Vandaar dus: biologische groenten.
De voordelen van het gebruik van de rode biet wegen ruimschoots op tegen de mogelijk optredende nadelen die door de bepaalde instantie onder de aandacht worden gebracht. Nadelen die niet optreden bij een gebruik zoals het hoort: bilogisch en niet mishandeld (fabrieksmatig bewerkt). Gewoon gebruiken zoals Cornelis Moerman dat aanbeveelt.
Een stabiel atoom lijkt op ons zonnestelsel, waarbij de elektrische lading van de kern (zon) in evenwicht is met de lading van de omringende elektronen (planeten). Ontbreekt er in de buitenste baan of schil een elektron, dan is het atoom instabiel. Een molecuul is een verbinding van meerdere atomen, waarvan de buitenste elektronen stabiele paren vormen. Een vrije radicaal is een atoom, al of niet als onderdeel van een molecuul, met een ongepaard elektron in de buitenste schil en is hierdoor instabiel. Het instabiele molecuul zal, in zijn streven naar evenwicht, een elektron van een ander molecuul afpakken, of aan een ander molecuul opdringen. Zo ontstaat een ketenreactie waarbij steeds meer vrije radicalen worden gevormd. Hoewel een vrije radicaal door elektronenoverdracht in evenwicht komt, is de functie van het betreffende molecuul blijvend beschadigd. Hierdoor worden celmembranen poreus, belangrijke enzymen onwerkzaam of kan zelfs de genetische informatie in de celkern worden veranderd. Dit leidt tot functieverlies en is de grondoorzaak van de symptomen van alle ziekten, veroudering, en degeneratie. Vreemd genoeg komen veel vrije radicalen voort uit de stof die essentieel is voor ons leven: zuurstof. Tijdens de energieproductie in onze cellen ontstaan veel zuurstofradicalen. Deze vrije radicalen, de singlet oxygen, het superoxide en de hydroxyl-radicaal, vormen de meest schadelijke en destructieve groep van radicalen in ons lichaam.
Andere veroorzakers van vrije radicaalschade hebben te maken met onze leefomgeving, voeding en gedrag. Bijvoorbeeld ioniserende straling zoals ultraviolet (zon)licht, elektromagnetische straling (elektra, magnetron, radar, zenders) evenals chemicaliën (zware metalen, oplosmiddelen, voedseladditieven), alcohol, roken, verkeerde voeding, stress, trauma en veroudering.
Toch zijn vrije radicalen niet altijd slecht. De fagocyten (vreetcellen) van ons immuunsysteem produceren vrije radicalen, waarmee ze bacteriën en andere indringers vernietigen. Dit zijn door enzymen gecontroleerde en gereguleerde radicaalreacties.
Een goede bescherming tegen de ongebreidelde en vernietigende kettingreacties van vrije radicalen is van levensbelang. Daarom hebben we in de loop van de evolutie een antioxidant systeem ontwikkeld. Antioxidanten kunnen één of meer elektronen afstaan om een radicaal te neutraliseren, zonder dat ze zelf een vrije radicaal worden. Zo stoppen ze de destructieve ketenreactie van radicaalvorming.
De spil van dit antioxidant-systeem is het enzym glutathion peroxidase, dat bovendien ook afvalstoffen en zware metalen uit ons lichaam afvoert. Als we jong en gezond zijn en via onze voeding voldoende grondstoffen ervoor binnenkrijgen, maakt ons lichaam per dag ongeveer 10 gram glutathion.
Voor een juiste werking van onze eigen (endogene) antioxidanten, is de aanvoer van antioxidanten via voeding (exogeen) noodzakelijk. Dit zijn o.a. de vitaminen A, C en E. Daarnaast zijn voor de aanmaak en recycling van onze antioxidant-enzymen voldoende vitaminen van het B-complex en de mineralen magnesium, selenium, zink, mangaan en koper noodzakelijk.
Het onschadelijk maken van vrije radicalen en de afvoer van gif- en afvalstoffen is de basis voor een optimale gezondheid en ziektepreventie.
Het lichaam produceert antioxidanten om de schade van vrije radicalen te beperken. Echter, onze moderne leefstijl, de toenemende milieuvervuiling en de verminderde aanmaak van antioxidanten tijdens het ouder worden (boven de 35 jaar) veroorzaken een tekort aan antioxidanten. Schade door vrije radicalen houdt verband met onder andere een verminderde afweer, voortijdige veroudering, hart- en vaatziekten, kanker en staar.
Ondersteuning van ons antioxidant systeem met behulp van supplementen bevordert de vitaliteit, versterkt het immuunsysteem en helpt verouderingsziekten te voorkomen.
Welke voeding en suppletie dient een vrouw te nemen, bij het gebruik van de anticonceptiepil?
De anticonceptiepil zorgt enerzijds voor een extra dagelijkse belastingvan het ontgiftingsproces dat via de lever verloopt en anderzijds voor eenverhoogd ziekterisico bij de vrouw door uitputting van belangrijke nutriënten zoals:
Vitamine C: is extra nodig om de lever te ondersteunen alsook wordt vitamine C sneller afgebroken door een verhoogde koper (ceruloplasmine) concentratie in het bloed ten gevolge van orale anticonceptie. Pilgebruiksters doen er dus goed aan hun dagelijkse menu te voorzien van verse groenten (rode paprika, broccoli, peterselie, kolen), groentesappen alsook vers fruit(acerola kers, rozenbottels, zwarte bessen en citrusvruchten) en nieuwe aardappelen die rijk zijn aan vitamine C.
Magnesium: zit vnl. in verse groene seizoensgroenten, ongeraffineerde granen, noten, rauwe melk, bruine rijst, garnalen en erwten. Bij een tekort kunnen gezondheidsklachten zoals vermoeidheid, duizeligheid, nervositeit, spierkrampen, hartkloppingen en hypoglycemie ontstaan.
Zink: een tekort aan dit mineraal maakt de pilgebruikster o.a. extra gevoelig voor hypoglycemie en infecties. Voorkom dit door elke dag minstens één van de volgende levensmiddelen te kiezen:volle graanproducten, oesters, schaaldieren, noten, vis, eieren, peulvruchten, rijst en bladgroenten.
Daarnaast is een goed multivitamine-mineralencomplex aangewezen om de dagelijkse tekorten door het pilgebruik aan te vullen: (volgens bloedanalyse vastgesteld)
Vitamine B1, B2, B3: 10-50 mg
Vitamine B5 en B6: 50-100 mg
Vitamine B12: 200-400 µg
Vitamine B9 of foliumzuur: 400 µg-2 mg
Inositol, choline: 50-75 mg
Taurine: 500-1000 mg
Vitamine C: 250-2000 mg
Vitamine E : 50-200 I.E.
Vitamine D: 1000 à 5000 I.E.
Magnesium: 300-500 mg
Kalium: 150-250 mg
Zink: 5-15 mg
Chroom: 100-200 µg
Mangaan: 3-5 mg
Ijzer (individueel te bepalen)
Pilgebruiksters dienen op te letten voor een verhoogd gehalte aan vitamine A, vitamine K en koper! Daarom is het aangewezen om een “rustperiode” van 3 tot 6 maand in te lassen tussen het stoppen van de pil en zwangerschap om dergelijke disbalans te kunnen herstellen.
Slechte darmen: de moeder van alle ziektes
Belangrijke aspecten maag-darm problematiek nader bekeken
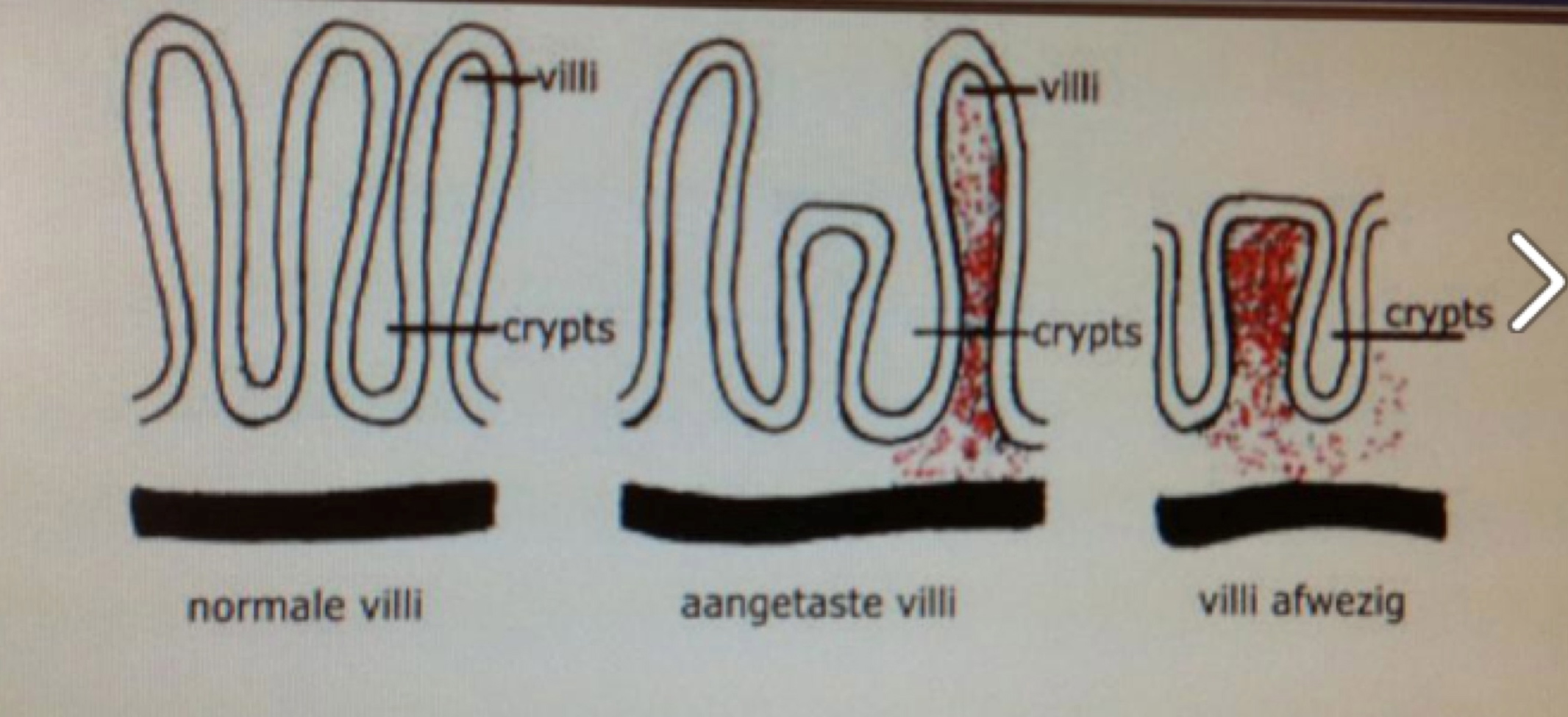
Voeding en vertering zijn onlosmakelijk verbonden met een goede gezondheid. Niet voor niets wordt gezegd dat we zijn wat we eten. Maar zelfs de meest gezonde voeding is pas doelmatig als die door ons lichaam goed kan worden opgenomen en verteerd. Een structureel slecht verteringsproces in maag en darmen kan leiden tot chronische klachten elders in het lichaam.
In India zegt men dan ook: ‘slechte darmen: de moeder van alle ziektes’.
Vage benaming maag-darmklachten geeft complexicteit aan
Klachten voortkomend uit het maag-darmkanaal zijn vaak moeilijk te omschrijven. Dat vinden we terug in de benaming. We praten over: spastische, lekkende of prikkelbare darmen. Benamingen die op zich weinig duidelijkheid scheppen.
Op de vraag wat een spastische of lekkende darm dan precies is, zal een eensluidend antwoord uitblijven. Het geeft aan dat er onduidelijkheid bestaat over de diversiteit aan oorzaken van de klachten. In deze publicatie zullen we ingaan op enkele belang-rijke aspecten van de maag-darmproblematiek:
Verstoord evenwicht van de micro-organismen
De zuurgraad in ons maag-darmkanaal
Weerstand, anti-lichamen en de rol van ons maag- darmstelsel
Darmen als onafhankelijk zenuwstelsel: ’the second brain’
Verstoord evenwicht micro-organismen
We spreken over een microbiële onbalans of dysbiose wanneer het evenwicht van de micro-organismen verstoord is. In deze publicatie betreft het de onbalans in het maag-darmtraject, de mond en de vagina.
In een situatie van microbiële onbalans kunnen bepaalde micro-ben ziektes veroorzaken doordat ze onze ingenomen voedingsstoffen ongunstig beïnvloeden en immuunreacties in het lichaam veroorzaken.
In onze darmen bevinden zich meer dan 500 verschillende soorten microben. Gezamenlijk ontwikkelen deze meer metabolische activiteit dan enig ander orgaan in ons lichaam.
Microben
De belangrijkste microben die bij een verstoord evenwicht ongewenst de overhand kunnen krijgen zijn:
parasieten
gisten en schimmels (vooral cadida albicans)
Virussen
bacteriën
Bacteriën kunnen verder worden onderverdeeld in:
* Aërobe bacteriën in het lichaam
Kunnen overheersen bij een overmaat aan antibiotica en bij acute virale of bacteriële infecties waardoor verschil lende goedaardige soorten worden gedood.
* Anaërobe bacteriën in het lichaam
De aanwezigheid van deze bacteriën in het lichaam is moeilijk vast te stellen.
* Bacteriën in voedsel en water
Zoals Salmonella, Shigella of E.coli die voedselvergiftiging kunnen veroorzaken.
De meest voorkomende bacteriën zijn: Campylobacter jejuni, Clostridium difficile en perfiringens, Bacteroides fragilis, Klebsiel-la, Proteus, Salmonella, Shigella, Cryptosporidiën.
Parasieten
Parasieten verdienen in het kader van de maag-darmproblematiek extra aandacht. Wereldwijd zijn meer dan 1 miljard mensen geïnfecteerd met parasieten. Mensen, dieren, voedsel en transportmiddelen zoals schepen zijn potentiële transporteurs van deze microben. Ziektes waaraan parasieten ten grondslag liggen nemen toe. Die toename heeft verschillende redenen:
* Sterke toename van internationaal reizen
* Immigratie
* Voedsel wordt tegenwoordig over de hele wereld gemaakt en getransporteerd
* Voedselbereiding in restaurants en de sterke toename van het “uit eten gaan”
* Toename wereldbevolking
Typische symptomen van een acute parasietinfectie in het beginstadium kunnen zijn, diarree, obstipatie, buikpijn, krampen, misselijkheid, eetlustverlies en een situatie waarin diaree en obstipatie elkaar afwisselen.
Meest voorkomende parasieten en hun klachten zijn:
Andere vaak voorkomende parasieten zijn:
Giardia lamblia, Cryptosporidia, Microsporidia, Dientamoeba fragilis.
Belangrijke vormen van microbiële onbalans
Veel essentiële functies van een gezonde darmflora zijn door de wetenschap inmiddels vastgelegd.
Een gezonde darmflora is o.a. afhankelijk van:
De beschikbaarheid van voldoende voedingsvezels
Gezond slijm
Een juiste zuurgraad
Goede doorbloeding
Gepubliceerd onderzoek geeft aan dat een verstoord evenwicht van de micro-organismen in maag en darmen de oorzaak is van, of bijdraagt aan: vitamine B12 tekort, prikkelbaar darmsyndroom, darmontstekingen, psoriasis, eczeem, acné en bepaalde auto-immuunreacties.
Belangrijke oorzaken van een verstoord microbieel evenwicht zijn:
Slechte voeding en nutriënten status
Stress
Het gebruik van antibiotica en bepaalde medicijnen
Verminderde darmperistaltiek
Maag-darm infectie
Xenobiotica (gifstoffen bv. door milieuvervuiling)
Verstoorde hormoonproductie (cortisol, insuline en schildklierhor monen)
Insuline resistentie
Hypochloorhydrie (weinig maagzuur)
Enkele belangrijke vormen van microbiële onbalans worden hier nader beschreven. Dysbiose door rotting
Dysbiose door rotting is een veel voorkomende vorm van microbiële onbalans. De belangrijkste oorzaak is het westerse voedingspatroon gekenmerkt door: veel vet, eiwit en vlees, weinig oplosbare vezels. Een dergelijk voedingspatroon veroorzaakt een verhoogde concentratie bacteroides. De stofwisseling verandert hierdoor zodanig dat het ingenomen voedsel in de darmen gaat rotten. De verhoogde concentratie bacteroides leidt tot een hogere activiteit van de enzymen beta-glucuronidase en urease. Deze enzymen kunnen galzuren omzetten naar toxines. Ook gaan ze de ontgifting van oestrogenen tegen, waardoor een verhoogd oestrogeengehalte in het lichaam kan ontstaan. Bovendien kunnen deze enzymen de zuurgraad (pH) in de ontlasting verhogen door een grotere ammonia productie.
Dysbiose door fermentatie
Dysbiose door fermentatie is een vorm van koolhydraat-intolerantie door een onbalans van microben, in het bijzonder van gisten en bacteriën die de koolhydraten in de dunne darm fermenteren. Dit kan leiden tot obstipatie, diarree, buikklachten, winderigheid en algehele malaise. Vooral mensen die zich niet prettig voelen na het eten of drinken van fruit, bier, wijn of bepaalde graanproducten kunnen te maken hebben met deze vorm van microbiële onbalans. Ook een teveel aan Candida albicans valt vaak in deze categorie.
Dysbiose door tekorten
Een dysbiose door tekorten ontstaat wanneer een persoon te weinig goedaardige darmbacteriën heeft zoals Lactobacillus en Bifidobacteria. Dit tekort kan bijvoorbeeld veroorzaakt worden door antibioticagebruik of te weinig vezels in de voeding. Mensen die deze vorm van microbiële onbalans hebben kunnen voedingsintoleranties krijgen en ook allergieën en infecties.
Dysbiose door overgevoeligheid
Bij een dysbiose door overgevoeligheid reageert het immuunsysteem zeer gevoelig op substanties in voeding en substanties geproduceerd door microben (endotoxinen). Mensen die last hebben van deze vorm van microbiële onbalans zijn multi-allergisch en hun symptomen zijn vaak systemisch; dus niet alleen in de darmen. Er is een grote kans dat mensen met auto-immuunziekten en mensen met een hoog IgA niveau te maken hebben met deze vorm van dysbiose.
De zuurgraad in ons maag-darm-kanaal
Bij een microbiële onbalans ontstaan door fermentatie, rotting en een hoge eiwitconsumptie diverse zure bijproducten. Omdat het lichaam zal proberen het zuur-base evenwicht te behouden raken hierdoor de alkali-reserves (basen) van onder andere de lever en alvleesklier uitgeput.
De zuren kunnen zich vanuit de darm een weg naar boven banen en zure oprispingen veroorzaken, die vaak ten onrechte worden aangezien voor een te grote maagzuurproductie. Deze zuren hebben geen verterend vermogen en werken zo de vertering van ons voedsel door maagzuur en de verteringsenzymen sterk tegen.
De darmen zijn veel gevoeliger voor het effect van de zuren dan de maag. In tegenstelling tot het dikke slijmvlies van de maagwand, dat is ingesteld op sterke zuren (maagzuur) is het dunne slijmvlies van de darmen dat absoluut niet.
Weerstand, antilichamen en ons maag-darmstelsel
In ons lichaam is het maag-darmkanaal het centrum van ons afweersysteem. Het bevat meer dan 80% van de cellen die antilichamen produceren.
De immuniteit die hier wordt opgebouwd is wezenlijk anders dan de opbouw elders in het lichaam.
Het maag-darm afweersysteem bestaat o.a. uit:
* Maagzuur- en verteringsenzymen
De eerste-lijnsdefensie tegen bacteriën en pathogenen.
* Het lymfeweefsel GALT
Het slijmvliesachtige oppervlak van de darmen is maar één cel dik. Daaronder bevindt zich het GALT (Gut Associated Lymphatic Tissue). Dit lymfeweefsel geeft antilichamen af.
* Anti-lichamen
Antilichamen openen de aanval op specifieke binnendringers (antigenen) zoals bacteriën, parasieten en voeding waarop allergisch wordt gereageerd. Tot de antilichamen behoren IgA, IgE, IgG, IgM, B cellen en het belangrijke SIgA.
Deze laatste antilichamen bespreken we nader. SigA eerste-lijns defensie
SIgA (Secretory Immunoglobuline A) zijn antistoffen die de eerste-lijnsdefensie in de darmen vormen.
Ze worden geproduceerd in speciaal hiervoor aanwezige cellen, Peyer’s plaque genaamd.
Deze cellen geven het SIgA af in de slijmvliesbarrière van darm.
SIgA is een belangrijk onderdeel van het afweersysteem in speeksel en in de slijmvliesmembranen van neus en longen. In gezonde darm-slijmvliezen is SIgA volop aanwezig. Het heeft daar als doel potentiële vijanden als bacteriën, schimmels en parasieten onschadelijk te maken. Dat geldt ook voor grote voedselstukken die ontstekingen kunnen veroorzaken. Door ontstekingsprocessen in de darm te voorkomen blijft de integriteit van de darmwand intact. Sommige mensen worden geboren met een lagere aanmaak van SIgA. Dit is meestal het geval bij mensen met allergieën en bij kinderen van ouders met allergieën. Zulke kinderen zijn meestal gevoeliger voor gluten, tarwe en voedselallergieën. Naast erfelijk bepaalde factoren zijn er ook andere elementen die invloed hebben op de hoeveelheid SIgA in ons lichaam.
We noemen er enkele:
Het hormoonniveau
Onze leeftijd: het SIgA niveau neemt met het ouder worden af
Vitamine A, zink, L-glutamine, probiotica en colostrum hebben de mogelijkheid om de productie van SIgA te verhogen
Stress
Voeding
Microflora van de darmen en de integriteit van de darmwand
Infecties
Borstvoeding
Andere soorten antilichamen zoals IgE, IgG en IgM zijn gespecialiseerd op specifieke indringers. Denk dan bijvoorbeeld aan voeding die allergie oproept of bepaalde soorten griepvirussen.
Een geheel andere manier van werken hebben lysozomen. Deze ondersteunen het afweersysteem in darmen, speeksel en tranen door de pathogenen ‘op te eten’.
Darmen als onafhankelijk zenuwstelsel: “the second brain”
De darmen hebben een complex onafhankelijk zenuwstelsel, ENS -Enteric Nervous System- genoemd. Deze ’tweede hersenen’ hebben een twee-weg communicatiesysteem met de hersenen. Dit verklaart de dramatische veranderingen die in het maag-darmkanaal plaatsvinden tijdens emotionele stress.
Zenuwprikkels -boodschappen-gaan vice versa van de hersenen naar de maag-darm over een weg die ‘vagus nerve’ wordt genoemd. Via dit ‘boodschappencircuit’ wordt de hersenen een signaal gegeven als er problemen zijn in het maag-darmtraject.
Om te kunnen communiceren gebruikt het ENS dezelfde neurotransmitters als de hersenen. In totaal zijn dit er ongeveer dertig, met serotonine als één van de voornaamste. Serotonine heeft ook een belangrijke rol bij de peristaltiek van de darmen evenals de zuurgraad aldaar.
In samenhang met de neurotransmitters kan ook obstipatie bezien worden. Slechts 25-35% van de mensen reageert bij obstipatie positief op een verandering van de voeding (bv. meer vezels). In de gevallen dat dit niet zo is moeten we de oorzaak dus elders zoeken. Het is dan goed om ons te realiseren dat ook een trage schildklierwerking of een beperkte darmperistaltiek tot obstipatie kunnen leiden.
Het is bekend dat serotonine een belangrijke rol speelt bij de peristaltiek. Beïnvloeding van de peristaltiek via ‘de tweede hersenen’ kan daarom in diverse situaties daarom succesvol zijn. Middelen die een positieve werking hebben op serotonine zijn anti-depressiva in een natuurlijke vorm.
Lekkende darm syndroom
Met het ‘lekkende darm syndroom’ wordt bedoeld: de conditie in de darmen waarbij de opname en het transport van voeding en voedingsstoffen verstoord is. Een belangrijke rol bij het lekkende darmsyndroom hebben de ‘villi’.
De villus is een vingerachtige structuur aan de oppervlakte van de dunne-darmwand (mucosa). Deze structuur vergroot de oppervlakte van de darm enorm en daarmee de opnamecapaciteit. De villus bestaat uit villi, microvilli en crypts.
Functies van de villi
* Villi zijn de werkpaarden van de darmen als het gaat om de opname van voeding in de bloedstroom
* Villi vermeerderen de oppervlakte van de dunne darm voor de absorptie van voeding
* Villi geven enzymen en bicarbonaat af ter vertering van voeding voordat deze kan worden opgenomen in de bloedbaan
* Villi absorberen het verteerde voedsel en transporteren dit naar de bloedstroom om zo verder in het lichaam gedistribueerd te worden
* Villi zijn een barrière waardoor bacteriën, parasieten en toxines worden geblokkeerd
Er zijn miljoenen microscopisch kleine villi in elke sectie van de dunne darm. Deze kunnen beschadigd raken zonder duidelijke symptomen. Villi zijn ontwikkeld om aangebrachte schade te compenseren. Wanneer echter grote secties villi zélf aangetast raken door ontstekingen, infecties of toxines dan heeft dit gevolgen voor de absorptie en het transport van voeding en voedingsstoffen. Dat geldt ook voor de enzym-afgifte en de defensieve mogelijkheden van villi. Zo kan o.a. het “lekkende darm syndroom ontstaan“.
Hoe ontstaat het lekkende darm syndroom
Voor het ontstaan van het lekkende darm syndroom zijn diverse redenen aan te wijzen. We bespreken enkele belangrijke oorzaken:
Voedingsallergie
Een voedingsallergie kan reactie in het darmweefsel veroorzaken die leidt tot verhoogde doorlaatbaarheid (permeabiliteit). Een lekkende darm laat dan de passage toe van abnormaal grote moleculen. Deze worden door het afweersysteem als vijandig (antigen) gezien zodat het deze aanvalt. Dit stimuleert het afweersysteem weer om méér antilichaamcomplexen te vormen wat tot een verslechtering van het darmweefsel leidt, de lekkende darm zal verergeren en zal meer allergische reacties veroorzaken. Als dit proces niet gestopt wordt kunnen de functies van lever, afweersysteem en darmen verslechteren.
Er is in het bijzonder een toename te zien van glutenintolerantie en -gevoeligheid en van de glutengerelateerde auto-immuunziekte Coeliakie.
Bacteriële onbalans
Wanneer een gezonde darmflora uit balans is geraakt door een overschot aan Candida albicans, gisten of parasieten spreken we van een bacteriële onbalans. Deze dysbiose kan leiden tot het lekkende darmsyndroom.
Ontstekingen
De immuunreacties (anti-lichaam respons) in de darmwand die normaal bedoeld zijn als protectie, kunnen ook juist ontstekingen creëren waardoor de lekkende darm verergert.
Malabsorptie voedingsstoffen
Wanneer aan de darmen door welke oorzaak dan ook voedingsstoffen worden onthouden kan binnen enkele dagen hyperpermeabiliteit ontstaan.
Genetische bepaling
Er blijken genetische patronen van hyperpermeabiliteit te bestaan. Zo zien we dat mensen met de ziekte van Crohn vaak al een verhoogde darmpermeabiliteit hebben. Maar ook familieleden van deze patiënten hebben vaak een verhoogde darmpermeabiliteit, ook al hebben ze zélf de ziekte van Crohn niet.
Chronische stress
Chronische stress vermindert de mogelijkheid van het afweersysteem om adequaat te reageren en gaat in het bijzonder het genezingspro-ces in de darmen tegen.
Ook de darmflora wordt door stress negatief beïnvloed.
Medicijngebruik
In het bijzonder antibiotica, farmaceutische hormonen (de pil, prednison e.d.) en N.S.A.I.D.’s (pijnstillers) kunnen een negatieve invloed hebben op de darmen.
Overige oorzaken
Overmatig alcoholgebruik, milieufactoren en slechte voeding. Toxische substanties in ongezonde darmen (zie dysbiose door rotting)
Toxische substanties in ongezonde darmen (zie dysbiose door rotting)
* Ammonia
Een bijproduct van ureum en eiwit decompositie, door bepaalde soorten microben in de darm. Ammonia wordt normaal omgezet naar ureum, als dit niet gebeurt kan het leiden tot neurologische symptomen en betrokkenheid bij foute transformatie van cellen.
* Histamine
Gevormd bij ontbinding van tryptofaan. Kan hoofdpijn, depressie, lage bloeddruk en misselijkheid geven.
* Indolen (Indican)
Gevormd door ontbinding van tryptofaan. Aantal indolen wordt vergroot door hoge vleesconsumptie.
* Mercaptan
Bijproduct van de ontbinding van cysteïne. Kan lage bloeddruk veroorzaken. Leidt tot stinkende winderigheid.
* Phenol
Bijproduct van de ontbinding van tyrosine in de dikke darm.
Kan necrose (weefseldood) van de darmmucosa en levercellen veroorzaken.
* Skatole
Bijproduct van de ontbinding van tryptofaan, gerelateerd aan anemie en malabsorptie syndroom. Skatole werkt acetylcholine en kalium tegen. Teveel skatole kan leiden tot slechte adem en stinkende ontlasting.
* Waterstofsulfiet gas
Bijproduct van eiwitontbinding.
Kan de wand van de darmen irriteren en kan even giftig zijn als cyanide. Het kan zwakheid, snelle polsslag en misselijkheid veroorzaken.
* Tyramine
Bijproduct van de ontbinding van tyrosine. Structureel verwant aan adrenaline. Kan tot hoge bloeddruk en problemen met het zenuwstelsel leiden wanneer het in het bloed circuleert.
Regelmatig eten van vis en noten zijn goed voor hart en bloedvaten,
(het immuunsysteem en de hersenen). Fibraten zijn reguliere cholesterol verlagers; LDL (slechte) daalt en HDL stijgt. Maar hebben ook bijwerkingen;
spierpijnen,
erectiestoornissen,
lever en galstoornissen en
ze verhogen de homocysteine spiegels.
Homocysteine zorgt weer voor verhoging voor hart en vaatziekten.
Statines verlagen de mortaliteit bij hart patiënten maar hebben ook bijwerkingen;
spierafbraak,
leverstoornissen,
slaap en geheugen stoornissen of
erectiestoornissen en
remmen de aanmaak van cholesterol. Cholesterol is een belangrijk onderdeel van de celwanden van al onze cellen, ze maken de celwand
soepeler, maar vooral in spiercellen en zenuwcellen. Statines zorgen dus voor starre celwanden, dus zwakte en krachtsverlies, geheugenproblemen en zenuwpijnen. Maw ouderdomskwaaltjes. Sporten verergert dit dus alleen maar.
Omega 3 vetzuren zijn goed voor hart en bloedvaten (DHA EPA) , kun je hypertriglyceridemie mee behandelen, komen het beste uit het onderzoek heeft geen bijwerkingen hebben eerder een meer voedend effect op vooral de cellen. Samenvatting van een artikel van pubmed: en andere artikelen
Alles in ons lichaam werkt alleen binnen bepaalde fysisch-chemische voorwaarden.
Dat zijn o.a. de juiste temperatuur, het juiste hormonaal evenwicht (daaronder horen alle lichaamshormonen), gebalanceerd immuunsysteem en
de juiste zuurgraad.
De juiste zuurgraad is dus een van de voorwaarden voor een goede gezondheid. Zonder deze is het niet mogelijk goed gezond te zijn! Alle celfuncties en dus alle orgaanfuncties gaan achteruit als de zuurgraad afwijkt. Geen enkel orgaan ontsnapt aan deze malaise. Bepaalde weefsels/functies zijn met meest gevoelig voor afwijkingen hierin, en dat zijn: botweefsel, haar, nagels, bloed(stolling), spieren, urinewegen, pezen en gewrichten. Een goede zuurgraad is voorwaarde voor behoud van een optimale gezondheid en voorwaarde voor behoud van met name gezonde botten.
Wat is het gevolg van verzuring?
Verzuring kan leiden tot botontkalking, doordat als het lichaam verzuurt, het gevolg is dat het lichaam dit compenseert, m.b.v. botzouten die alkalisch zijn. Dit alles onder invloed van het parathormoon, het hormoon van de bijschildklier. Er vindt niet alleen demineralisering van het bot plaats, maar vaak ook van haren en nagels.
Behalve botontkalking kan verzuring leiden tot nierstenen. Zure urine geeft namelijk een verschuiving van opgelost calciumoxalaat en urinezuur naar de kristalvorm. Er treedt dus kristallisatie op. Dat is zijn microscopisch kleine kristallen. Daarna ontstaat niergruis en daarna nierstenen.
Hetzelfde principe vindt plaats in lichaamsweefsels. Bij een hoge zuurgraad vind kristallisatie plaats van de bufferzouten. Deze slaan neer in de weefsels. Het gevolg is verharding, verstijving van ligamenten, gewrichtskapsels.
Onder invloed van verzuring gaat de werking van het immuunsysteem veranderen. Er onstaat een verhoogde neiging tot ontsteking. Verder bestaat een concrete invloed van de zuurgraad op de vertering. Je verteringsenzymen werken bij een bepaalde zuugraad. Als deze afwijkt gaat de vertering achteruit.<
Verzuring leidt dus tot:
1. verstijving van spieren, pezen, gewrichtskapsels, 2.Demineralisering van botten, nagels en haren, m.a.g. botontkalking, nierstenen, verteringsproblemen
Eerst een nog paar extra opmerkingen, die je moet begrijpen en om je te motiveren zodat je zuurgraad als een van de basisvoorwaarden voor je gezondheid gaat zien.
Als we ouder worden gaat ons lichaam geleidelijk aan verzuren. Een deel van de veroudering ontstaat door verzuring. Je weefsels gaan verharden omdat er onder invloed van verzuring o.a. zure (buffer)zouten in neerslaan. Dit geeft o.a. een verharding van bindweefsel en dus ook verstijving. De stijfheid die bij het ouder worden optreedt is deels hiervan een gevolg. De verzuring leidt tot een overreactie van bepaalde immuuncellen met als gevolg brand! Dit kan uitmonden in een soort veenbrand. En dit is oorzaak van heel veel ellende. Alle biochemische reacties in ons lichaam verlopen met behulp van enzymen. Enzymen functioneren optimaal bij een bepaalde zuurgraad. Wijkt de zuurgraad af dan wijkt de werking van deze enzymen af.
Eiwitten in de voeding geven verzuring. Maar eiwitten hebben we nodig. Dus moet er in de voeding compensatie zitten voor de verzurende eiwitten bijvoorbeeld meer groente en fruit consumeren. Evolutie
Tijdens de evolutie is ons genenpakket ontstaan in samenhang met de omgeving waarin de vroege mens leefde en in samenhang met vooral het voedsel dat voorradig was. Het originele voedsel van de mens was heel rijk aan groente en fruit, knollen, zaden, noten. Af en toe at de mens vermoedelijk vlees en of vis. De mens is een opportunist door zijn intelligentie en eet wat hij kan verkrijgen. Granen en brood, pasta’s, rijst at de vroege mens niet, het was er eenvoudigweg niet. Dit oerdieet zullen de meeste van ons niet erg aantrekkelijk meer vinden. Het was wel goed! Relatief weinig snelle koolhydraten, weinig verzadigd vet en niet al te veel eiwitten en daardoor niet verzurend. We zullen de voeding en biochemie van de oermens moeten nabootsen. In onze moderne maatschappij kunnen we niet leven als de oermens. We zullen op een andere manier dezelfde biochemie moeten bereiken.
Kan ik met gezonde voeding mijn zuurgraad in orde houden? Het is uit te rekenen wat je moet eten om niet te verzuren. Dat komt voor de meerderheid van ons neer op een pond tot een kilo groente en fruit per dag. Omdat dit niet voor ieder realiseerbaar is moeten we kunstgrepen toepassen. Kunstgrepen zijn in dit verband voedingssupplementen. Wat moet je doen om de juiste zuurgraad te bereiken? Stap 1. Eerst bepalen of er sprake is van verzuring.
Methode: met behulp van PH strips, kun je de zuurgraad van je urine meten.
Dit moet je op een doorsnee dag alle keren doen dat je plast. De volgende uitslagen zijn mogelijk A. Ochtendurine, dit is de eerst urine van de dag, d.w.z. de eerst na 06.00 in de ochtend, niet eten/drinken ( water mag wel) na 20.00 uur vorige avond Er zijn nu 2 mogelijkheden.
1. Deze heeft een PH lager dan 6,0
2. Deze heeft een PH hoger dan 6,0
B. Dag urine; alle keren dat je overdag plast de zuurgraad meten
Er zijn nu 3 mogelijkheden
De urine PH is overdag alle keren beneden de 7,0.
De urine PH komt 1 x op 7,0 of hoger.
De urine PH is overdag meerdere keren boven de 7,0
Mogelijkheden:
1. ochtendurine lager dan 6,0 en dagurine alle keren onder de 7,
2. ochtendurine lager dan 6,0 en dagurine 1 x 7,0 of hoger
3. ochtendurine lager dan 6,0 en dagurine meerder keren 7,0 of hoger
4. ochtendurine hoger dan 6,0 en dagurine alle keren onder de 7,0<
5. ochtendurine hoger dan 6,0 en dagurine 1 x 7,0 of hoger
6. ochtendurine hoger dan 6,0 en dagurine meerdere keren hoger dan 7,0
Mogelijkheid 6: heel goed, goede zuurgraad
Mogelijkheid 5: redelijk tot goed
Mogelijkheid 4: matig
Mogelijkheid 3: goed
Mogelijkheid 2: matig tot redelijk
Mogelijkheid 1: slecht
Stap 2. Nu je zuurgraad bekend is kun je bepalen wat je moet doen,
Let op: gebruik je medicijnen roep dan deskundige hulp in! Je kunt nog steeds ontzuren, maar dat kun je beter onder begeleiding doen! Mogelijkheid 3,5,6: je doet het voor wat uw zuurgraad betreft goed genoeg. Je hoeft geen maatregelen te nemen. Mogelijkheid 2: je doet het matig tot redelijk. Het is voldoende meer groente en fruit te eten. Mogelijkheid 1,4: onvoldoende.
Advies: eet (veel) meer groente en fruit. Neem daarbij supplementen om je zuurgraad verder te verbeteren. neem daarbij contact met me op. Stap 3. Check of je voldoende maatregelen hebt genomen
Hoe weet je of je het voldoende doet? Of de maatregelen afdoende zijn?
Doe nog een keer de test als hierboven en kijk of je in een betere categorie komt.
LET OP Check nadat je een paar dagen supplementen gebruikt je urine. De PH mag niet hoger worden dan 7,5.
Stap 4. Houdt deze maatregelen, die je in categorie 1 hebben gebracht dan een minimaal ½ jaar vol. Controleer af en toe je urine en zie erop toe dat de PH niet boven de 7,5 komt. Als dat wel gebeurt, neem dan minder supplementen. Je lichaam heeft tijd nodig om te ontzuren, om zijn overtollige zuren kwijt te raken, vooral als deze al zijn uitgekristalliseerd.
Stress is een biologische en psychologische reactie die we ervaren op het ontmoeten van een bedreiging en dat gevoel hebben we dan ook.
Een stressor is de stimulus (of dreiging) dat stress veroorzaakt, bijvoorbeeld examen, scheiding, overlijden van dierbare, verhuizing, verlies van werk.
Plotselinge en ernstige stress produceert over het algemeen
Verhoging van de hartslag
Toename van de ademhaling (longen verwijden)
Afname van de spijsvertering activiteit (geen honger)
Lever zorgt er voor dat glucose wordt vrijgegeven voor energie
Ten eerste, ons lichaam beslist als rechter of een situatie stressvol is. Deze beslissing is genomen op basis van zintuiglijke input en verwerking (dat wil zeggen de dingen die we zien en horen in de situatie) en ook op de opgeslagen geheugens (dat wil zeggen wat er gebeurd is de laatste keer waren we in een soortgelijke situatie).
Als de situatie beoordeeld wordt als zijnde stress, dan wordt de hypothalamus (aan de basis van de hersenen) geactiveerd.
De hypothalamus in de hersenen is verantwoordelijk voor de stressrespons. Wanneer een stressreactie wordt geactiveerd, worden er signalen uitgezonden naar twee andere structuren:
de hypofyse en het
bijniermerg.
Deze korte termijn antwoorden worden geproduceerd door The vecht-of vluchtreactie via de Sympathomedullary Pathway (SAM). Lange termijn stress wordt gereguleerd door de hypothalamus hypofyse-bijnier (HPA)-systeem.
De hypothalamus hypofyse-bijnier (HPA)-systeem
De stressor activeert de hypothalamus hypofyse as
De hypothalamus stimuleert de hypofyse
De hypofyse scheidt adrenocorticotroop hormoon (ACTH)
ACTH stimuleert de bijnieren om het hormoon corticosteroïden te produceren.
Cortisol stelt het lichaam in stand de gestage aanvoer van bloedsuiker te houden
Adequate en stabiele bloedsuikerspiegel te helpen persoon om te gaan met langdurige stress, en helpt het lichaam om terug te keren naar de normale
De bijnierschors geeft stresshormonen cortisol. Deze hebben een aantal functies, waaronder los opgeslagen glucose in de lever (voor energie) en controle zwelling na letsel. Het immuunsysteem wordt onderdrukt, terwijl dit gebeurt.
Sympathomedullary Pathway (SAM)
De hypothalamus activeert ook het bijniermerg. Het bijniermerg is onderdeel van het autonome zenuwstelsel (ANS).
De ANS is het deel van het perifere zenuwstelsel die fungeert als een besturingssysteem, het handhaven van homeostase in het lichaam. Deze activiteiten worden over het algemeen uitgevoerd zonder bewuste controle.
Het bijniermerg scheidt het hormoon adrenaline. Dit hormoon zorgt ervoor dat het lichaam klaar gestoomd wordt voor een vecht-of vluchtreactie. Fysiologische reactie inclusief een verhoogde hartslag.
Adrenaline leidt tot het opwekken van het sympathische zenuwstelsel en verminderde activiteit in het parasympatische zenuwstelsel.
Adrenaline maakt veranderingen in het lichaam, zoals afname (bij de spijsvertering) en verhoogt (zweten, verhoogde hartslag en bloeddruk).
Zodra de ‘dreiging’ over is, neemt de parasympathische tak de controle over en brengt het lichaam terug in een evenwichtige staat.













 hansstoop
hansstoop